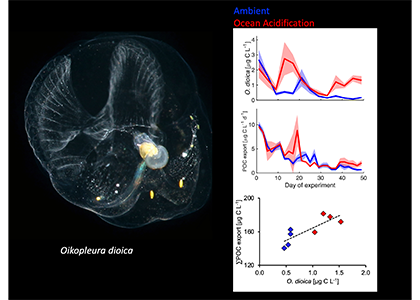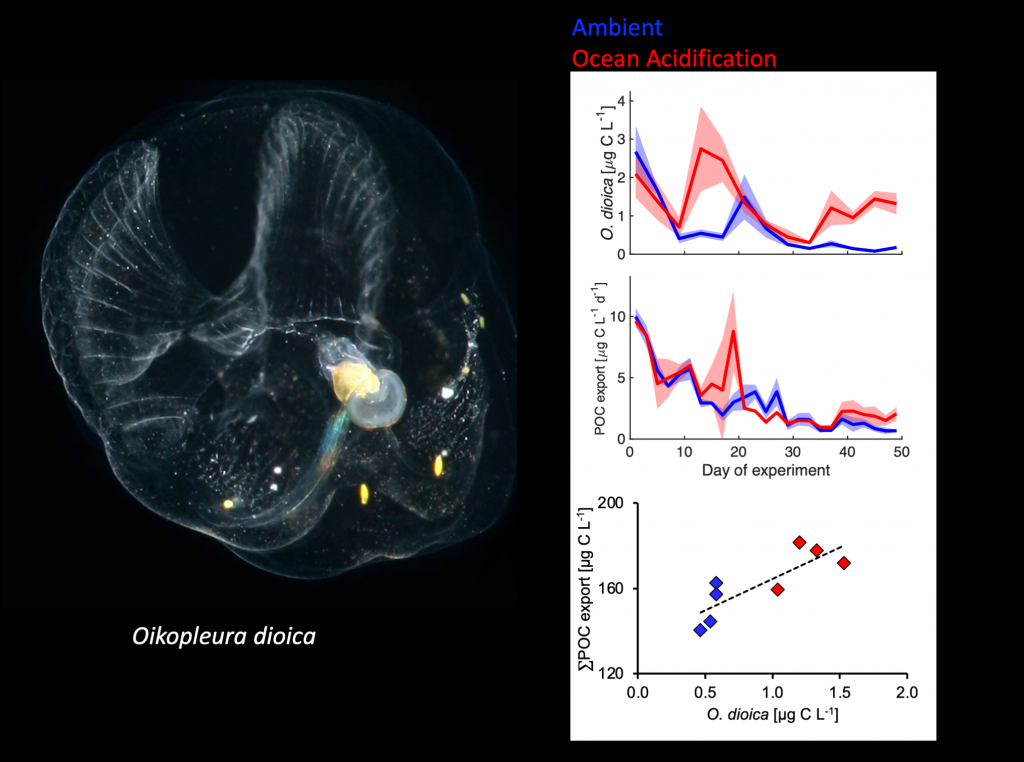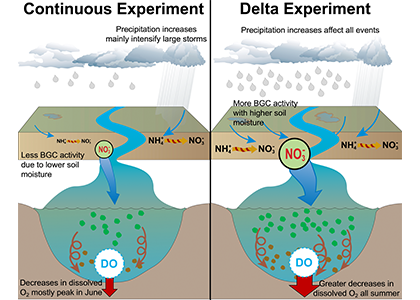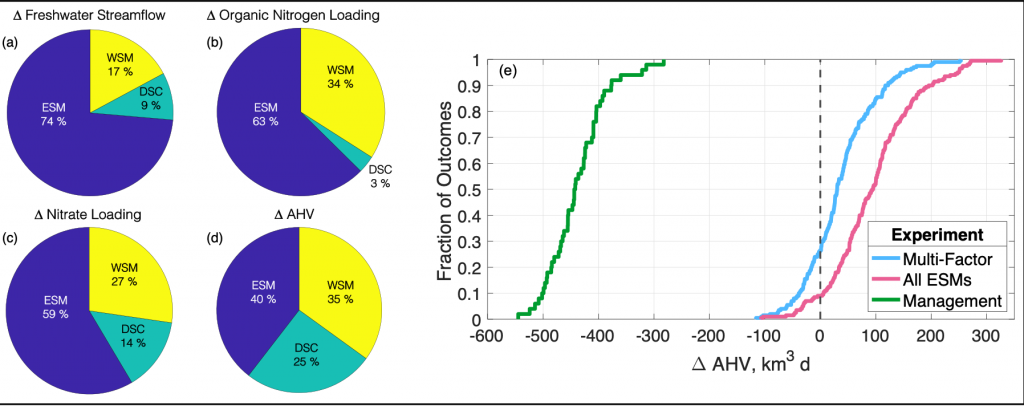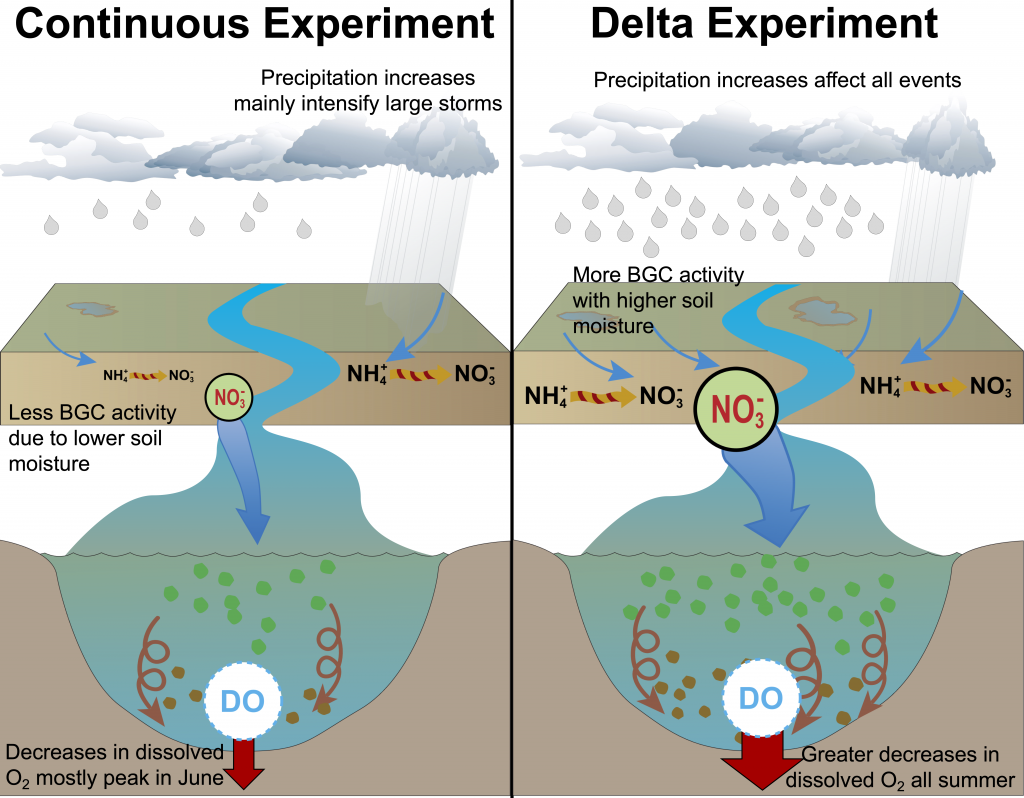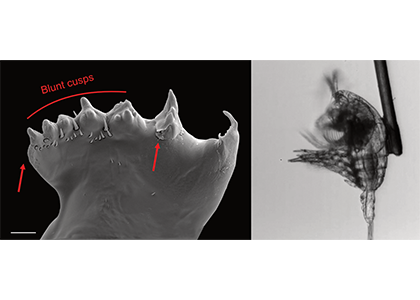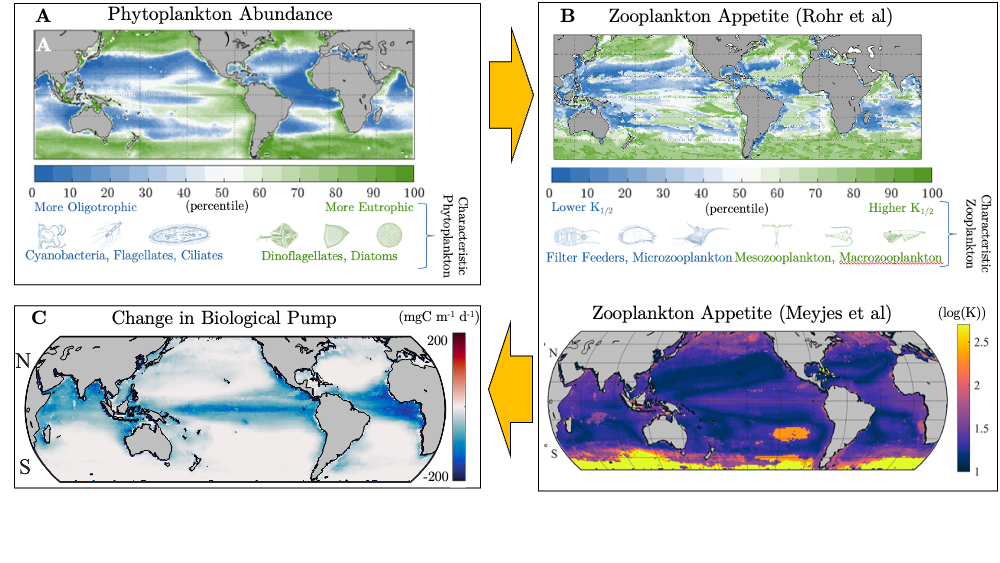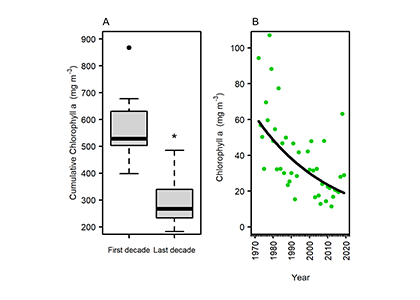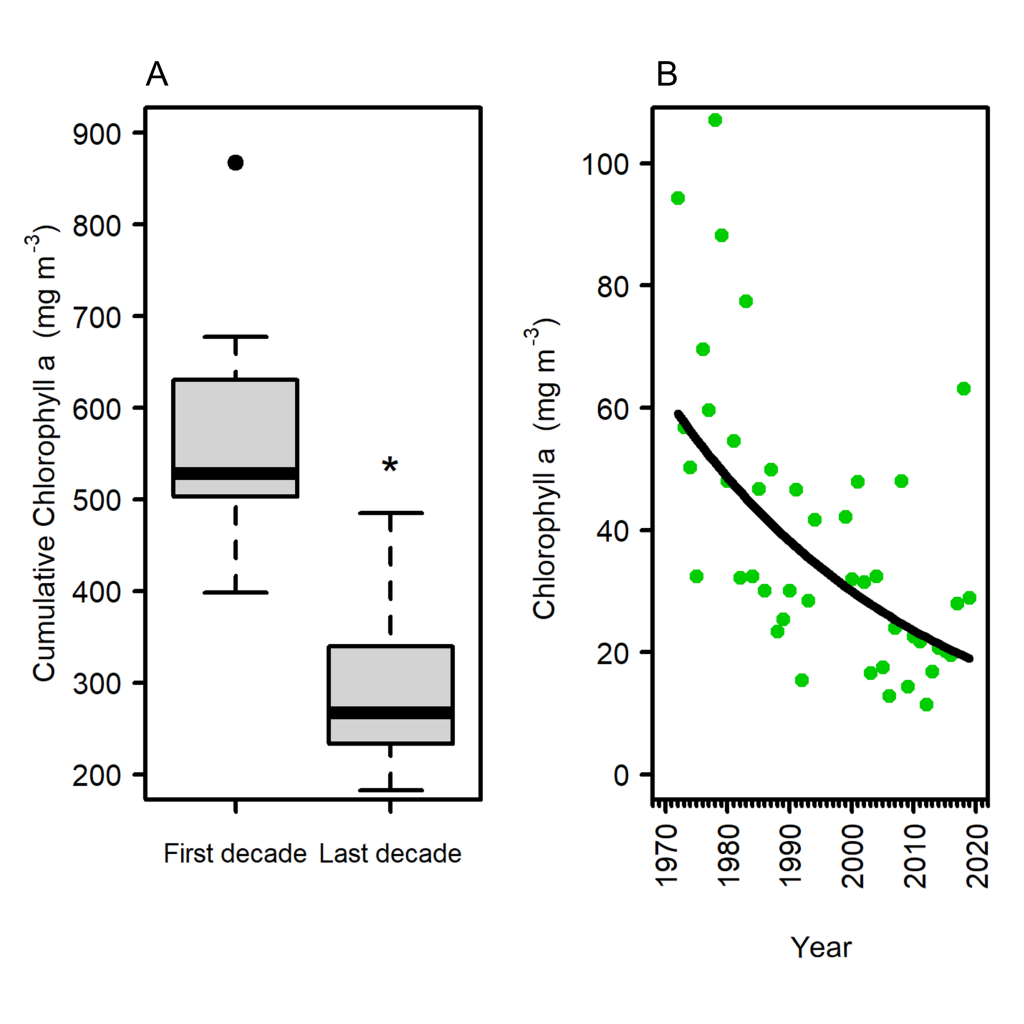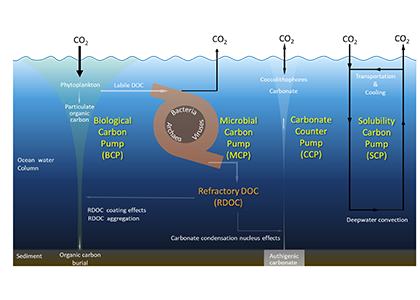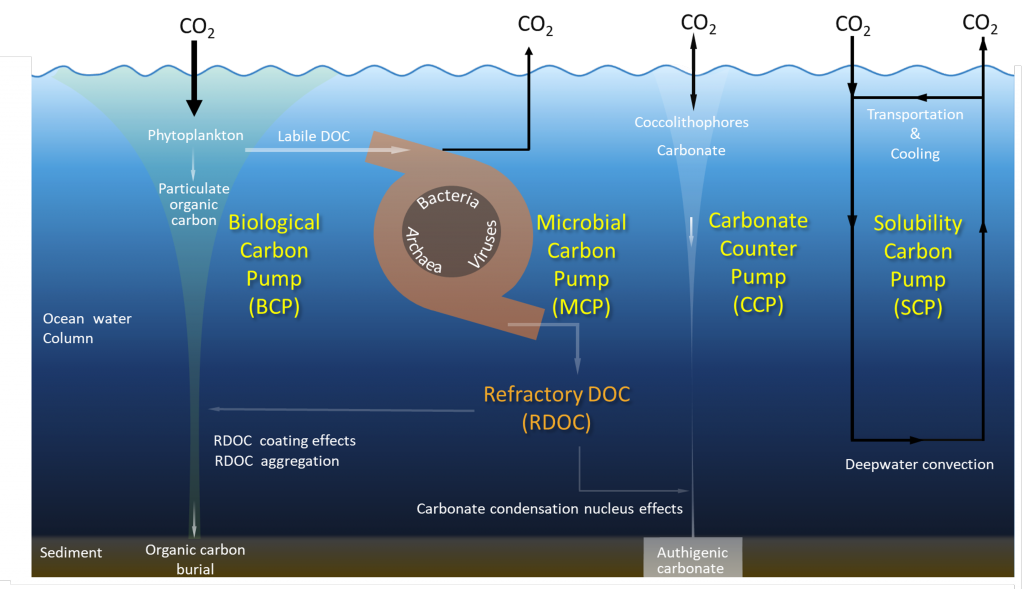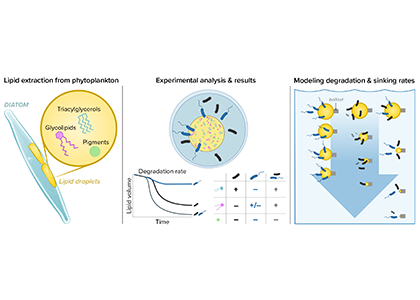
What drives carbon from the atmosphere to the deep ocean? The journey of phytoplankton-derived carbon is critical in the global carbon cycle, yet the influence of interacting bacteria in degrading lipid-rich particles during their descent has remained a mystery—until now.
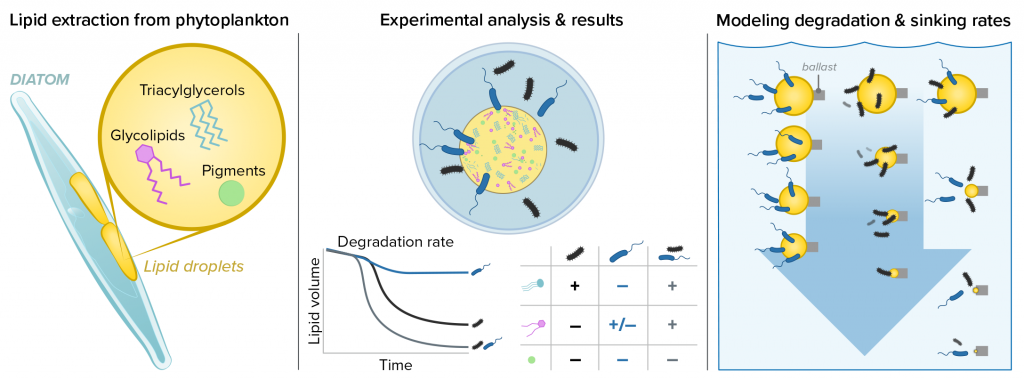
Using an innovative combination of nano-scale lipidomics and microscopy, researchers investigated how bacteria target and degrade diverse lipid molecules in sinking oceanic particles. The study, published in Science, revealed that bacteria exhibit distinct dietary preferences, governed by their lipid-degrading genes rather than taxonomic affiliation. Interactions among bacteria influenced both degradation rates and timing, together reshaping our view on the efficiency of lipid transport to the ocean depths. These findings were incorporated into a mathematical model, revealing how microbial communities could regulate the carbon transfer efficiency.
This research enhances our understanding of the ocean’s carbon pump, highlighting the pivotal role of bacterial communities in carbon sequestration. By uncovering how microbial interactions affect carbon transfer, these findings improve climate models and support the development of strategies to mitigate atmospheric CO2.
Authors
Lars Behrendt (Uppsala University, Sweden)
Benjamin van Mooy (Woods Hole Oceanographic Institution)
Twitter: LarsBehrendt4
Bluesky: @belab1.bsky.social