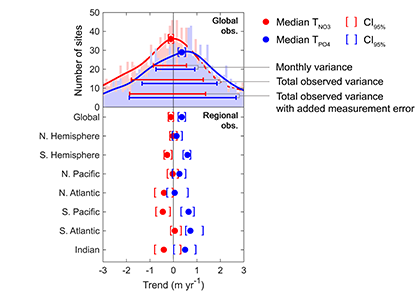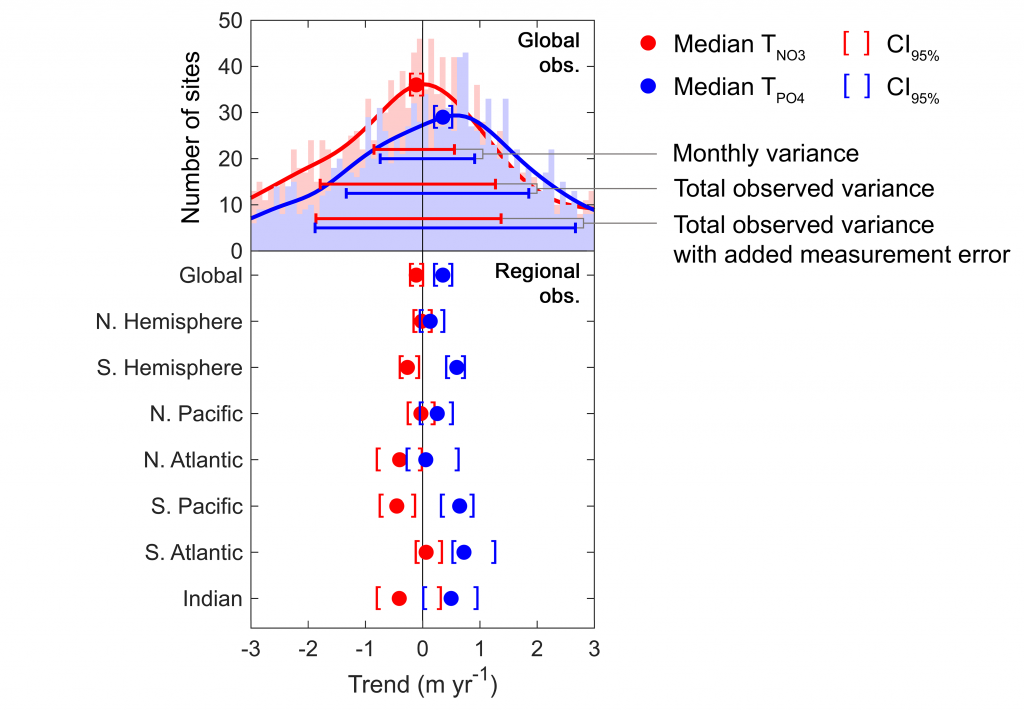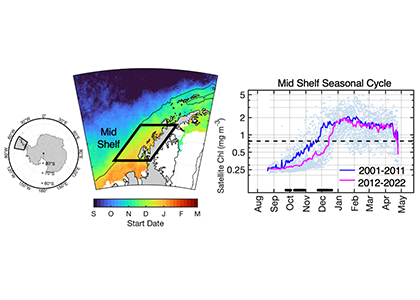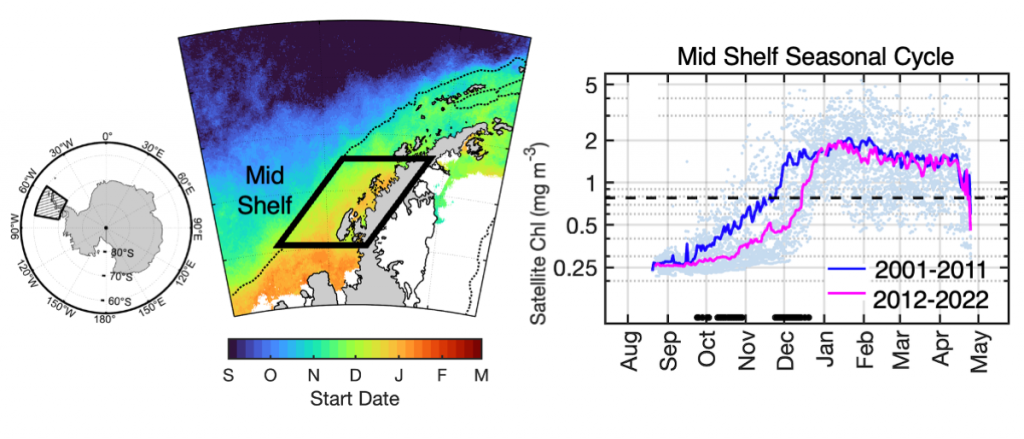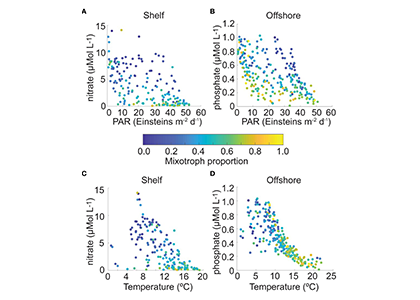
How do phytoplankton respond to short-term changes in temperature and nutrient availability, and how does this response vary under different baseline conditions?
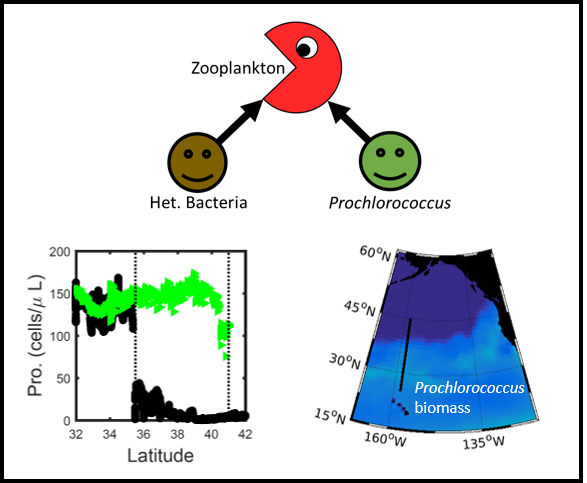
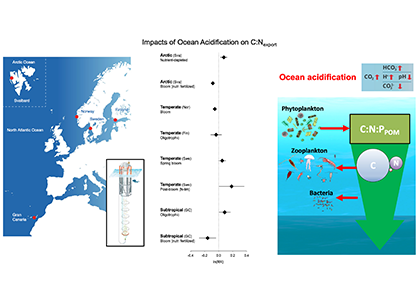
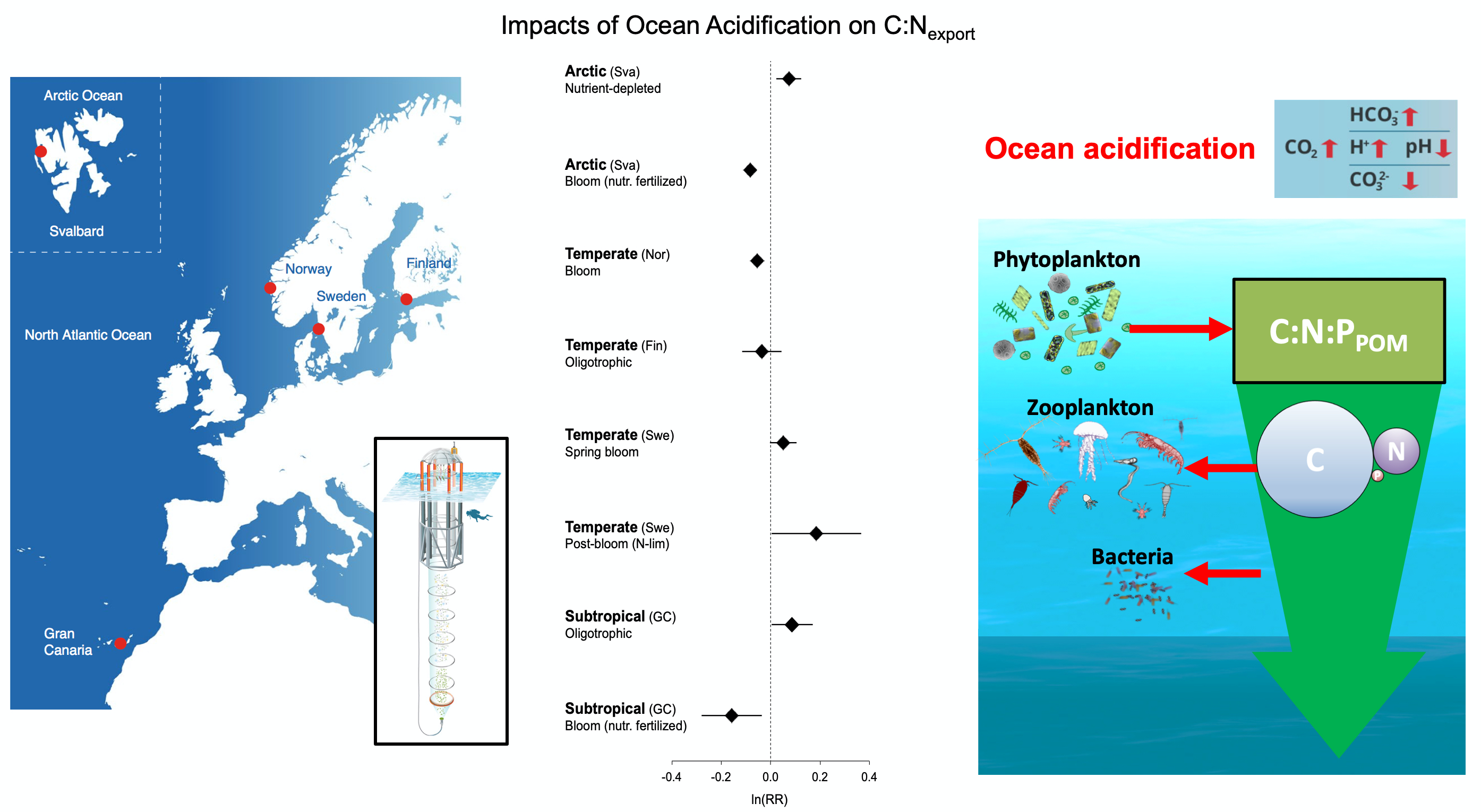
A recent study perturbed a summer plankton community by manipulating nutrient concentrations and temperature (±4°C) in short-term controlled microcosms to assess the relative impacts of each factor on phytoplankton physiology and community structure. Nutrient amendments rapidly shifted the phytoplankton community towards larger cells, altered elemental stoichiometry, and reduced microzooplankton grazing. Conversely, temperature changed community composition, but had no impact on size or stoichiometry. These results paralleled a similar spring incubation and regional field studies, which together suggest that over short time scales (days) nutrient amendments may produce more consistent and significant impacts on phytoplankton composition and physiology than temperature or grazing. Though temperature is a significant driver of phytoplankton dynamics globally, the impacts of short-term temperature changes, like marine heat waves, may depend on baseline thermal conditions, the starting plankton community, and the availability of nutrients to cope with increased metabolic demands. Understanding baseline conditions will be central to deciphering how short-term temperature changes may impact phytoplankton communities and the ecosystems they support.
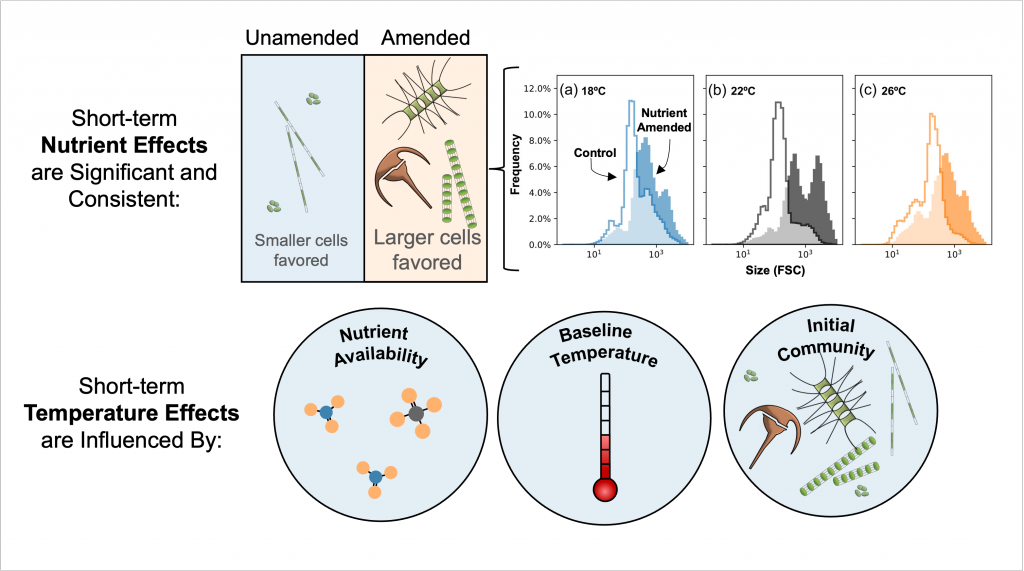
Figure caption. Illustrative diagram showing the impacts of short-term nutrient amendments on phytoplankton communities, and the factors that influence the thermal response. Frequency distribution plots show how the community size structure <20 µm differed between the nutrient amended treatment (solid) and the control (white) on day 4 of the incubation experiments, as discerned via flow cytometry forward scatter (FSC).
Authors
Stephanie I. Anderson (University of Rhode Island)
Gayantonia Franzè (University of Rhode Island & Institute of Marine Research)
Joshua D. Kling (University of Southern California)
Paul Wilburn (Michigan State University)
Colin T. Kremer (University of Connecticut)
Susanne Menden-Deuer (University of Rhode Island)
Elena Litchman (Michigan State University)
David A. Hutchins (University of Southern California)
Tatiana A. Rynearson (University of Rhode Island)
Bluesky
@drplankton.bsky.social
@quant-ecology.bsky.social
@elenalitchman.bsky.social
@stephanieianderson.bsky.social