
The presence of large-scale Ekman downwelling is the textbook explanation for low nutrient concentrations, and hence low productivity, in subtropical gyres. However, recent research has suggested that mesoscale eddies oppose and substantially reduce this downwelling, a process known as eddy cancellation (Doddridge et al, 2016). Eddy cancellation represents a substantial alteration to the widely accepted notion of large-scale Ekman downwelling in subtropical gyres, and motivates our study of the processes that determine nutrient concentration within subtropical gyres.
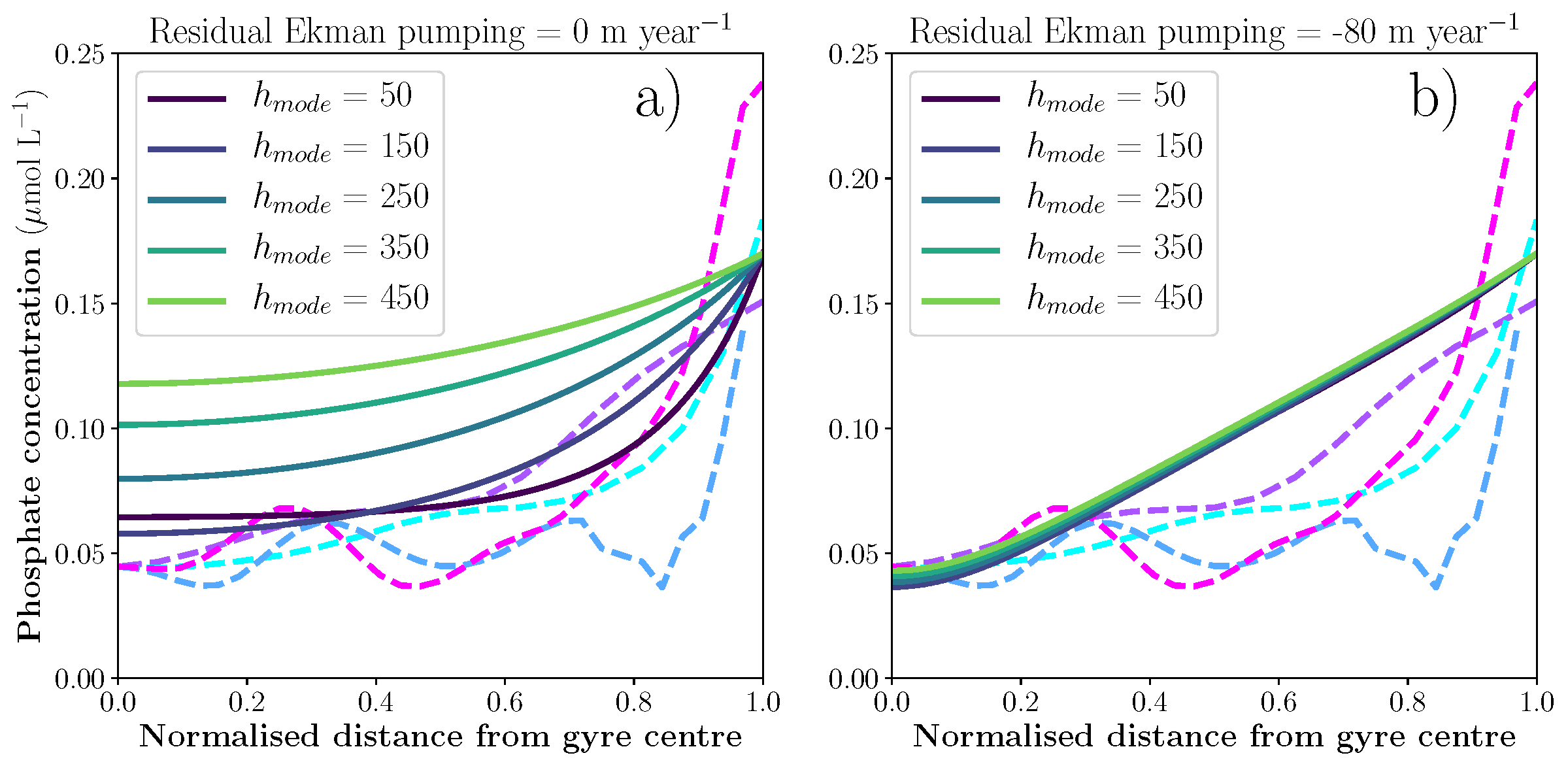
Figure 1: Sensitivity experiments for mode water thickness (hmode) with two values of residual Ekman pumping. a) With no residual Ekman pumping, phosphate concentration responds strongly to mode water thickness. b) When Ekman pumping is strong, phosphate concentration does not depend on mode water thickness. The dashed lines represent transects of climatological phosphate concentration in the euphotic zone of the North Atlantic subtropical gyre (Garcia et al., 2013).
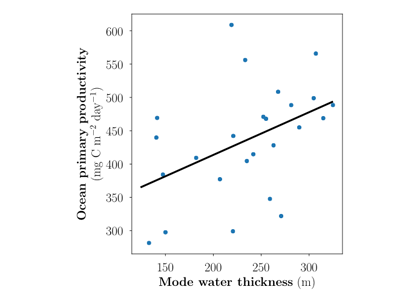
A recent paper published in the Journal of Geophysical Research: Oceans and featured in an MIT News article describes an idealized model for nutrient concentration in subtropical gyres that can account for this reduction in Ekman pumping. The model predicts that surface productivity is sensitive to the thickness of the underlying subtropical mode water layer, provided that the residual Ekman pumping is small (Figure 1). Comparison of this prediction with observations from the Bermuda Atlantic Time series Study (BATS) shows that surface productivity increases as the thickness of the underlying mode water increases (Figure 2), as predicted by the idealized model in the absence of substantial Ekman pumping.
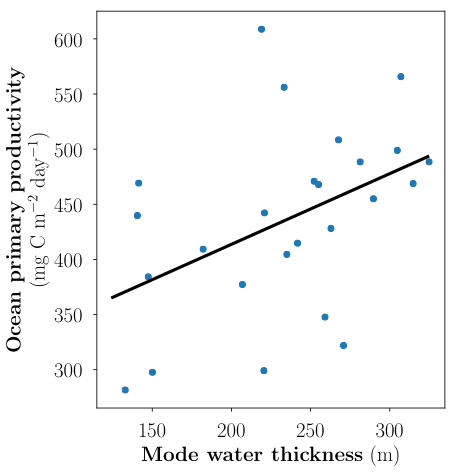
Figure 2: Annually averaged primary productivity and mode water thickness from the BATS dataset. The linear fit between mode water thickness and primary productivity is statistically significant (p ≈ 0.027) and explains 19.5% of the variance in primary productivity.
The observed relationship between productivity and mode water thickness at BATS is consistent with a small residual Ekman pumping, indicating highly effective eddy cancellation in the subtropical North Atlantic. Previous research (Palter et al., 2005) has suggested that as the subtropical mode water layer thickens, it blocks nutrient entrainment from below, resulting in lower productivity in the euphotic zone. However, this study suggests that a thicker subtropical mode water layer actually increases the surface nutrient concentrations by promoting more effective recycling of nutrients within the gyre. With a thicker mode water layer, more of the nutrients in the particulate flux are remineralized before they pass through the thermocline and become isolated from the surface ocean. This means that a thicker mode water layer leads to higher nutrient concentrations and supports primary productivity in subtropical gyres. This represents a fundamental change in our understanding of how nutrients are supplied to the surface waters of subtropical gyres.
Authors:
Edward Doddridge (Department of Earth, Atmospheric and Planetary Sciences, MIT)
David Marshall (Atmospheric, Oceanic & Planetary Physics, University of Oxford)
See the Eos spotlight on this research

