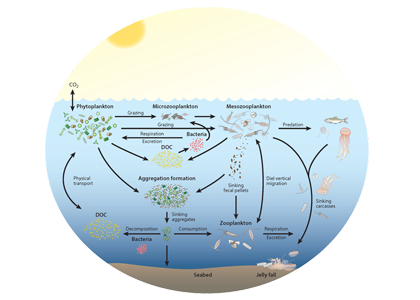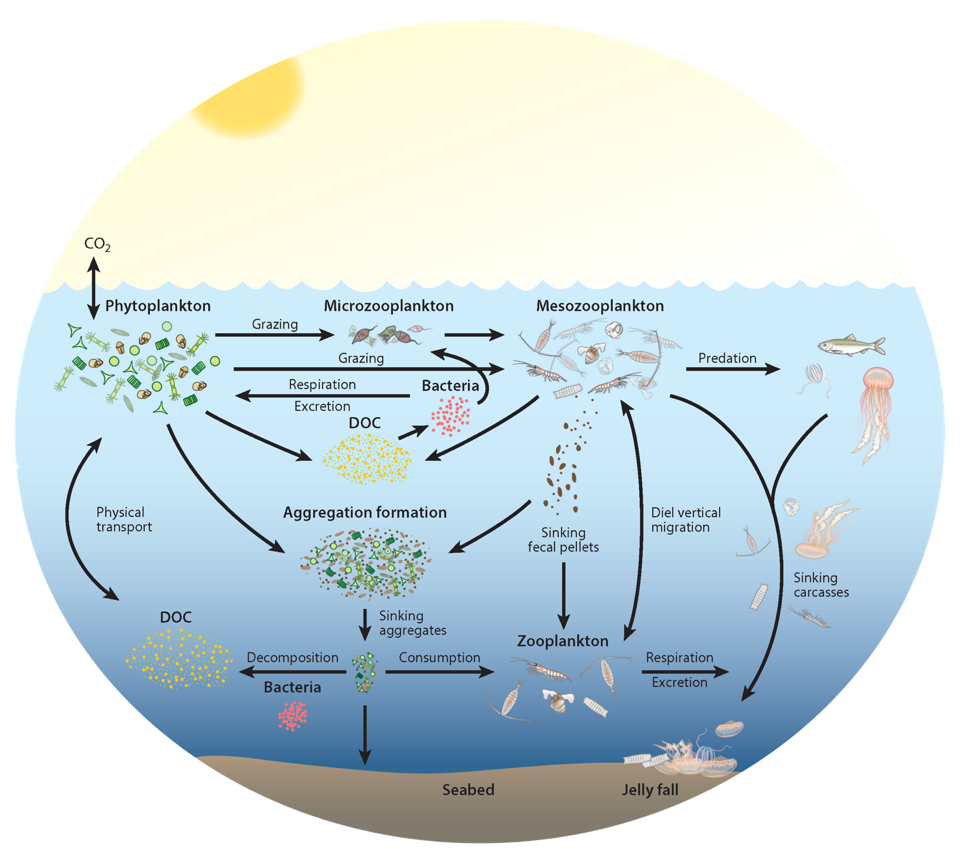
The removal of bioavailable nitrogen (N) by anaerobic microbes in the ocean’s oxygen deficient zones (ODZs) is thought to vary over time primarily as a result of climate impacts on ocean circulation and primary production. However, a recent study in PNAS using a data-constrained model of the microbial ecosystem in the world’s largest ODZ revealed that internal species oscillations cause local- to basin-scale fluctuations in the rate of N loss, even in a completely stable physical environment. Such ecosystem oscillations have been hypothesized for nearly a century in idealized models, but are rarely shown to persist in a three-dimensional ocean circulation model.
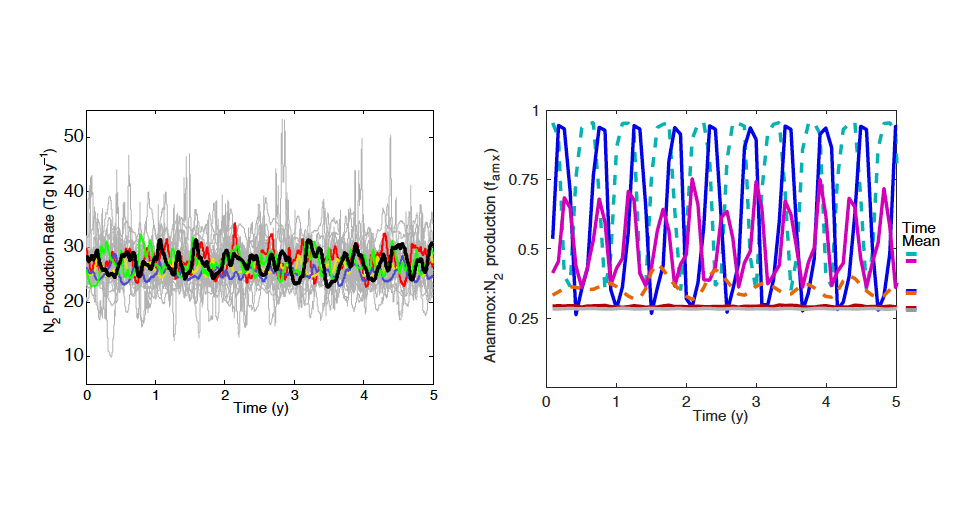
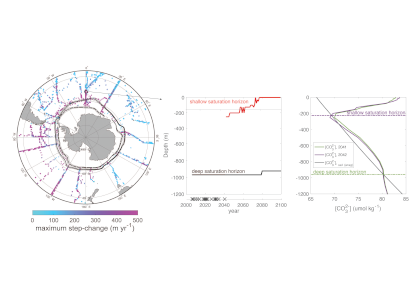
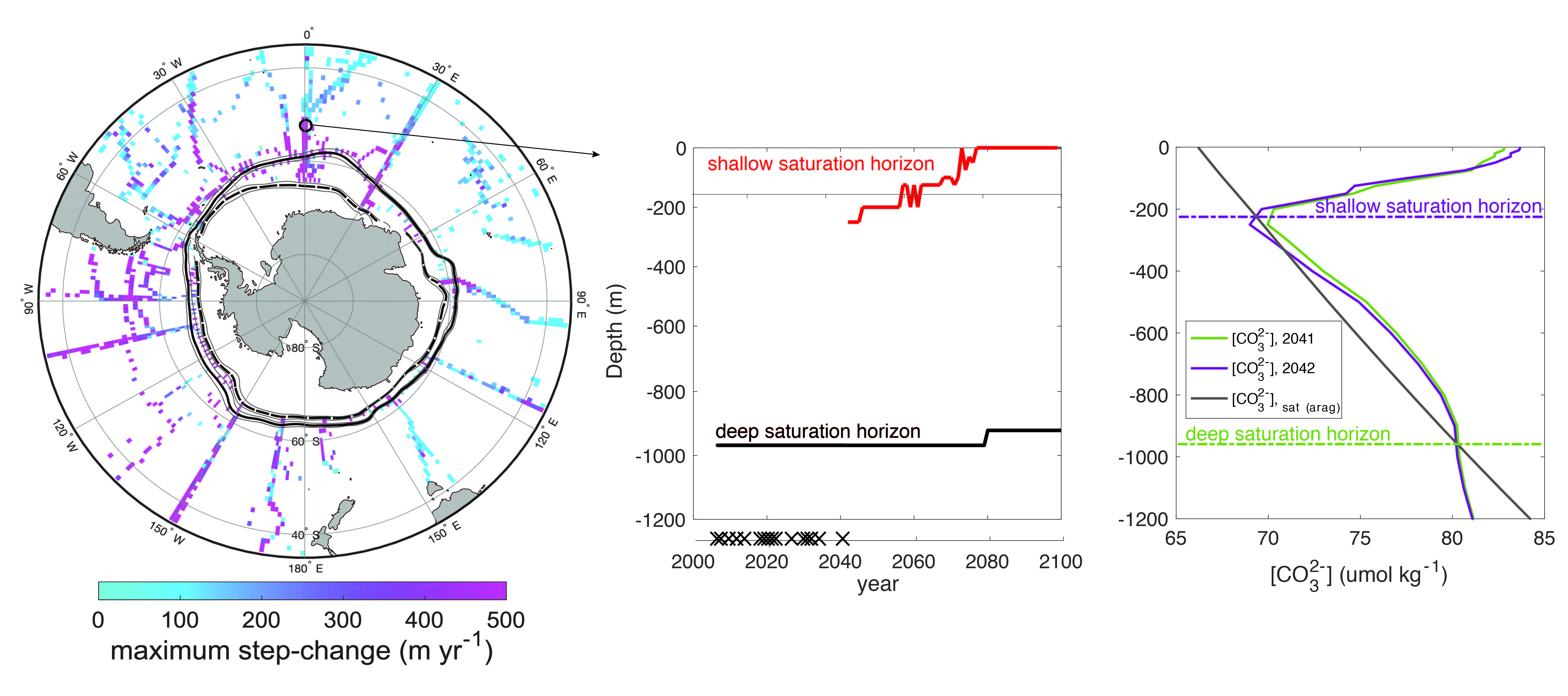
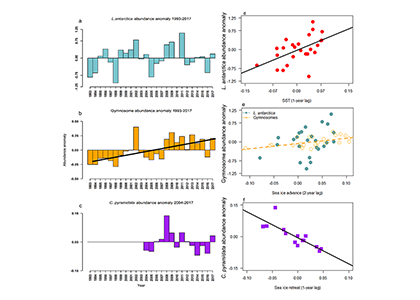
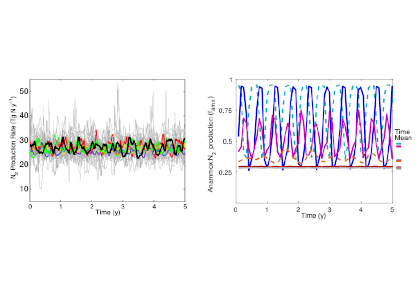
Figure caption. Ecological variability in the basin-scale rate of nitrogen loss over time (left) and in the local-scale contribution of autotrophic anammox to total N loss (right) in a model with unchanging ocean circulation. In the left panel, colors represent model simulations with different biological parameters. In the right panel, colors represent distinct locations within the ODZ in the standard model simulation.
These emergent ecosystem dynamics arise at the oxic-anoxic interface from O2-dependent resource competition between aerobic and anaerobic microbes, and leave a unique geochemical fingerprint: infrequent spikes in ammonium that are observable in nutrient measurements from the ODZ. Non-equilibrium ecosystem behavior driven by competition among aerobic nitrifiers, anaerobic denitrifiers, and anammox bacteria also generates fluctuations in the balance of autotrophic versus heterotrophic N loss pathways that help reconcile conflicting field observations.
These internally driven fluctuations in microbial community structure partially obscure a direct correspondence between the chemical environment and microbial rates, a universal assumption in biogeochemical models. Because of the fundamental nature of the underlying mechanism, similar dynamics are hypothesized to occur across wide-ranging microbial communities in diverse habitats.
Authors:
Justin L. Penn (University of Washington)
Thomas Weber (University of Rochester),
Bonnie X. Chang (University of Washington, NOAA)
Curtis Deutsch (University of Washington)
See also the OCB2019 plenary session: Anthropogenic changes in ocean oxygen: Coastal and open ocean perspectives (Monday, June 24, 2019)