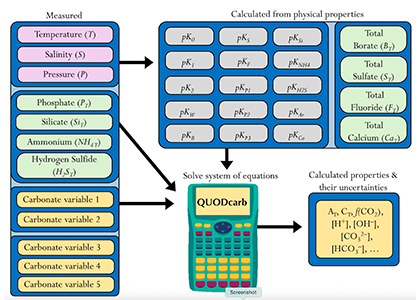
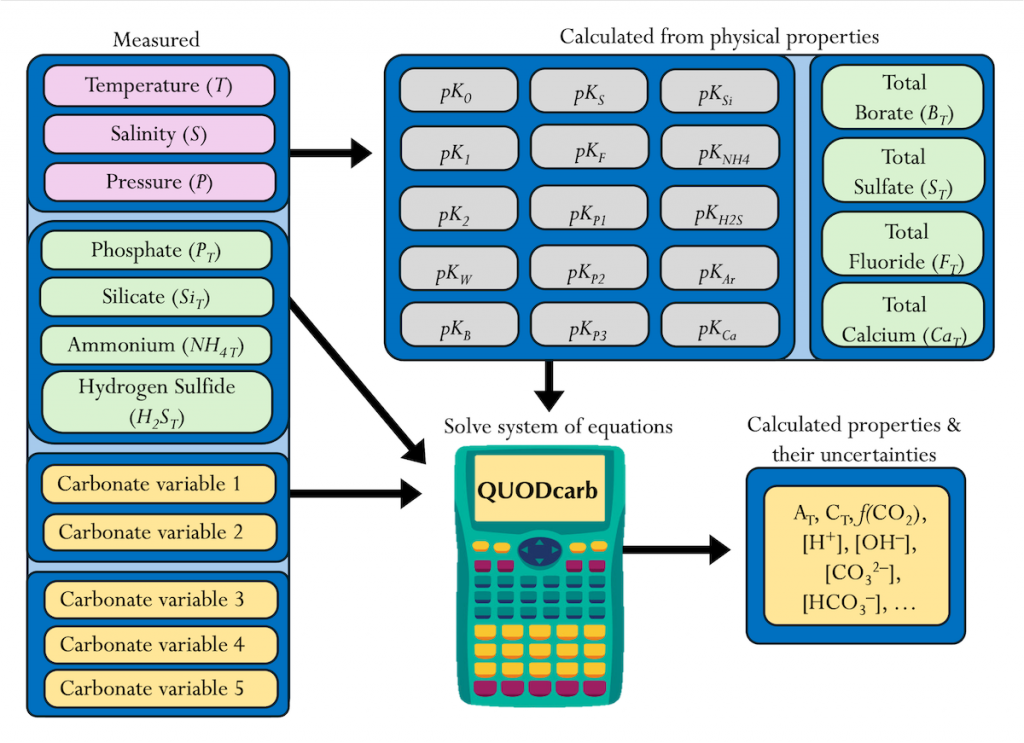
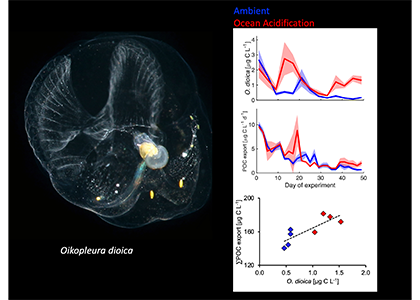
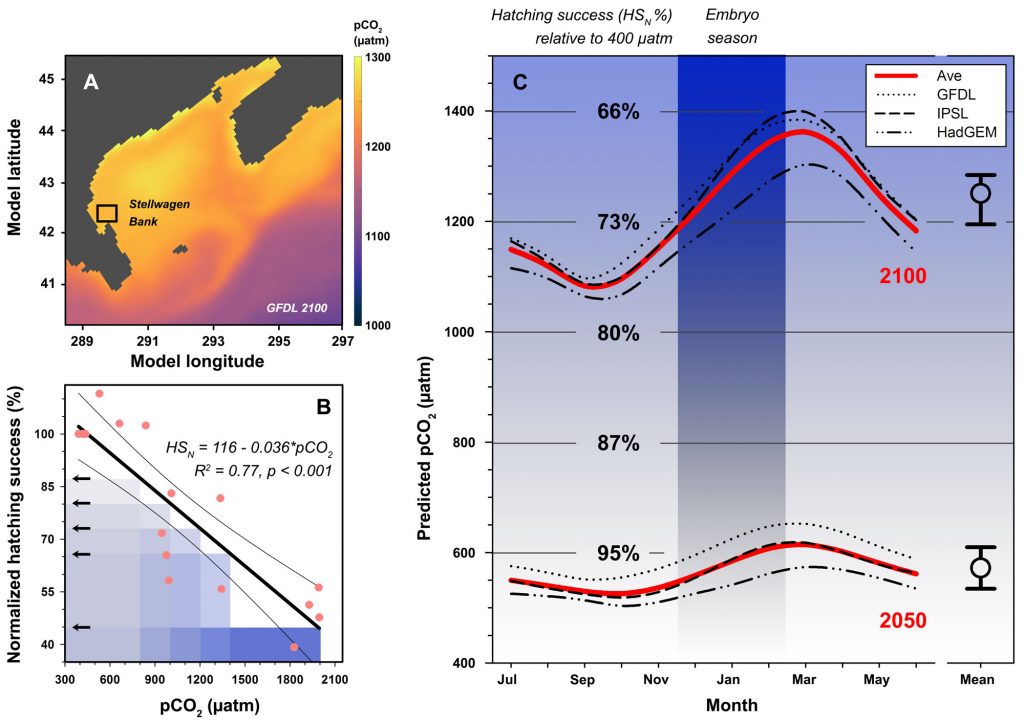
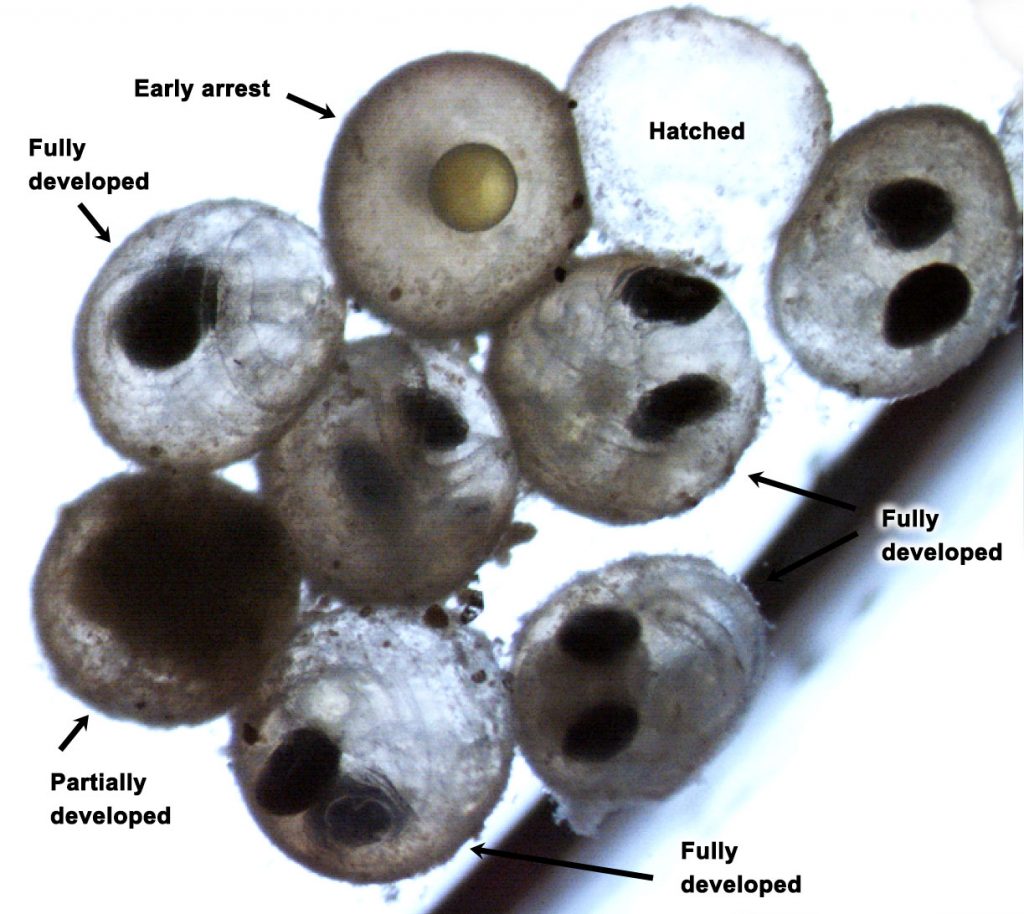
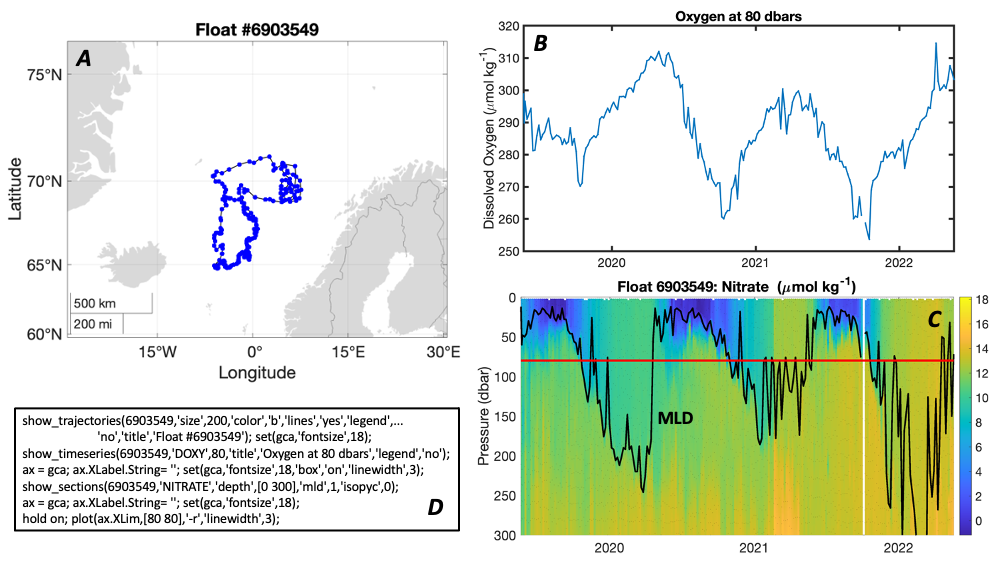
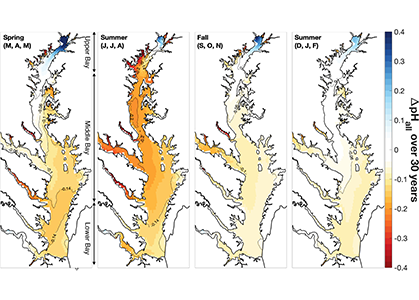
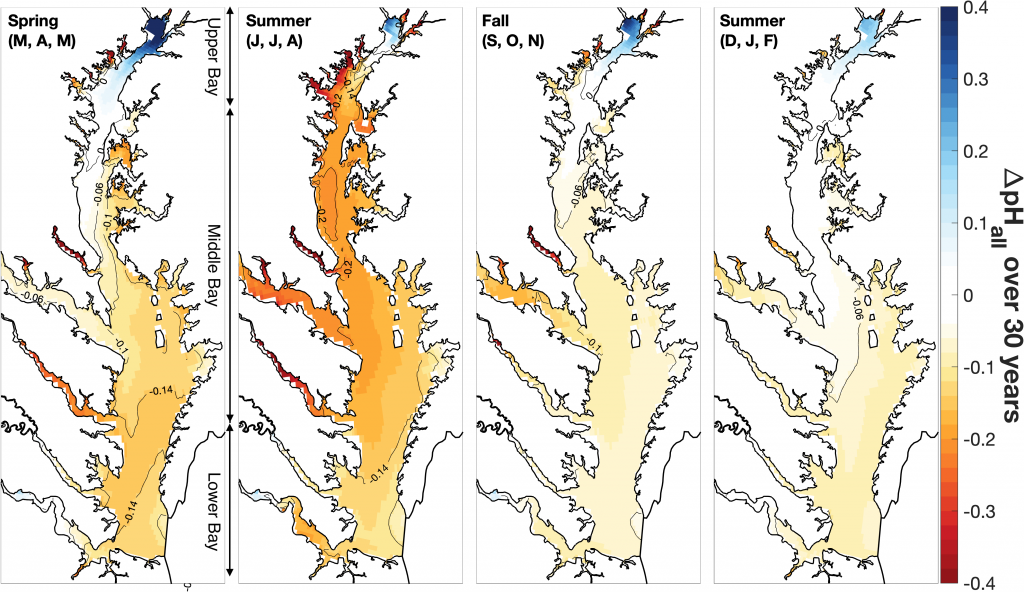
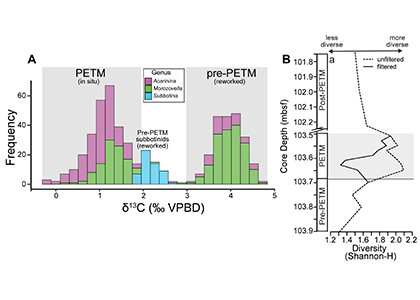
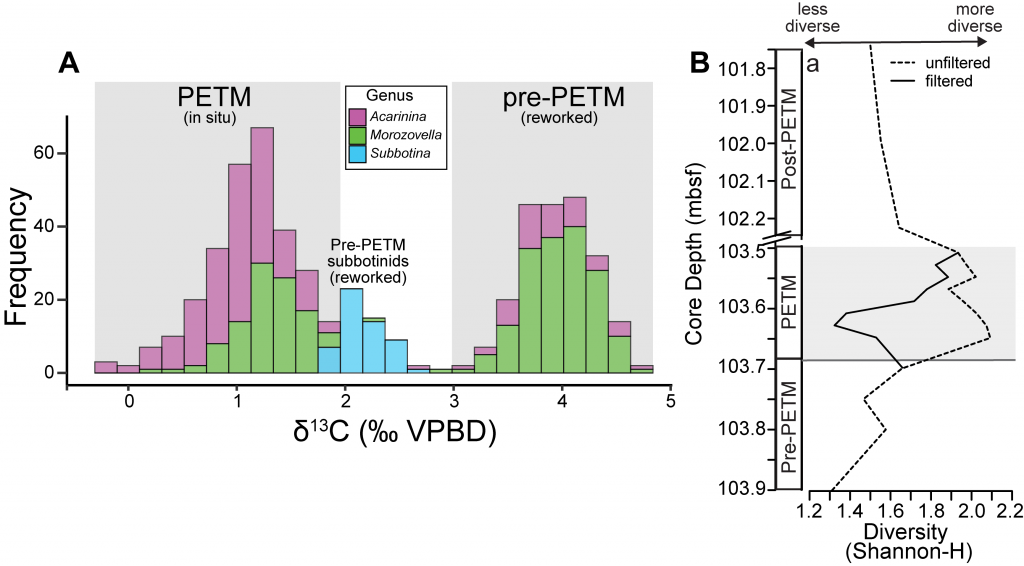
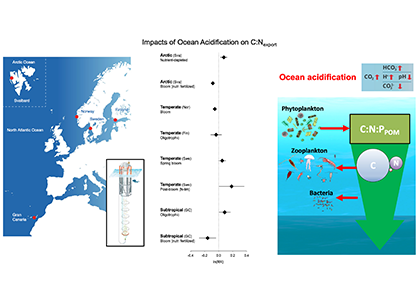
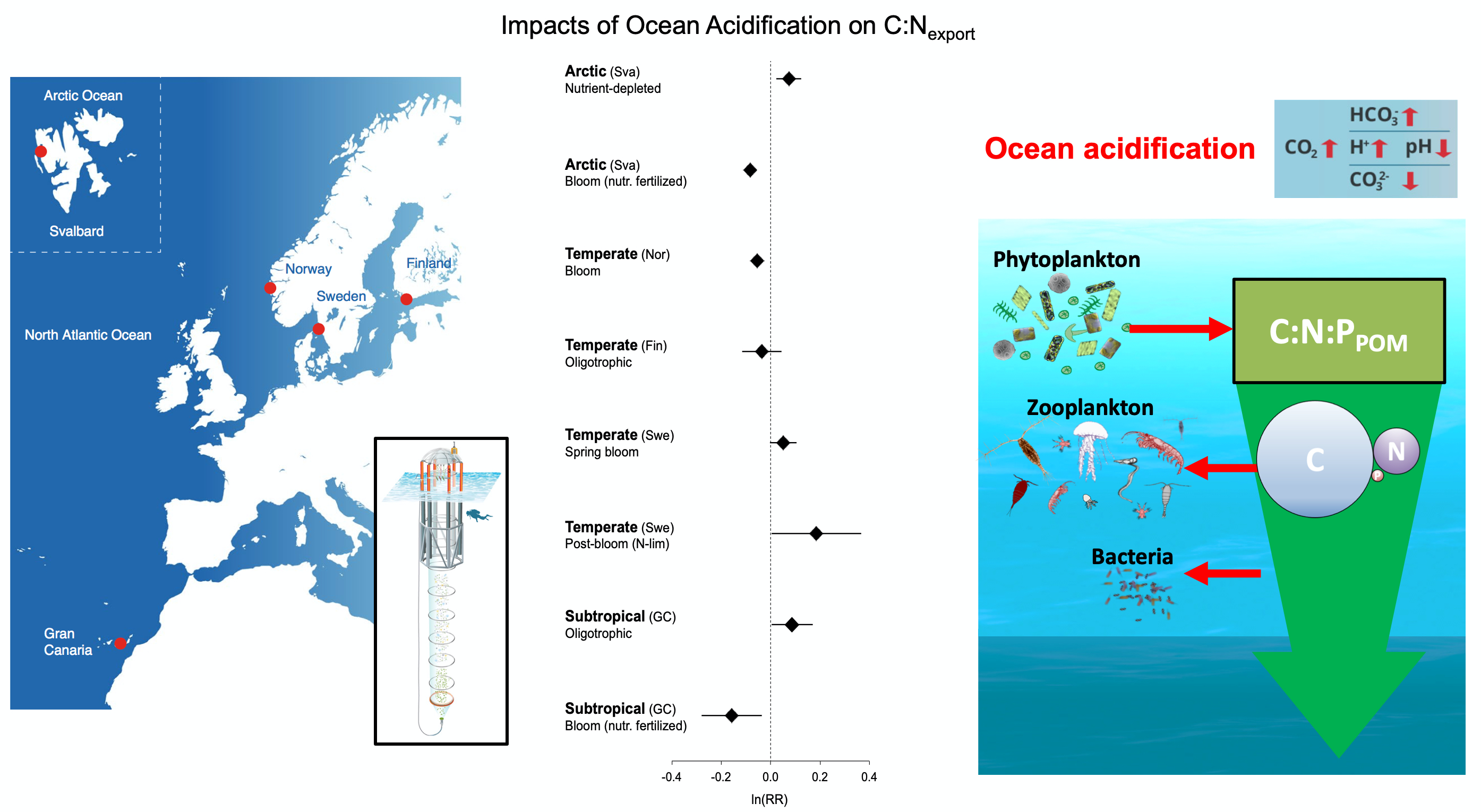
The paper provides a comprehensive synthesis of 68 existing ocean carbonate chemistry data products and data product sets, including cruise-based compilations, time-series datasets, gap-filled observational products, and model-based reconstructions. The authors highlight the diversity of available products, noting differences in spatial coverage, temporal resolution, methodologies, and intended scientific applications. By systematically cataloguing and comparing these datasets, the study helps researchers identify which products are most suitable for specific scientific questions related to ocean carbon cycling and ocean acidification.
Interface for the most updated list of products

Authors
Li-Qing Jiang (University of Maryland; NOAA National Centers for Environmental Information; Scripps Institution of Oceanography)
Amanda Fay (Columbia University / Lamont-Doherty Earth Observatory)
Jens Daniel Müller (ETH Zürich; Carbon to Sea Initiative)
Luke Gregor (ETH Zürich; Swiss Data Science Center)
Alizée Roobaert (Flanders Marine Institute, VLIZ)
Lydia Keppler (Vycarb Inc.)
Dustin Carroll (Moss Landing Marine Laboratories; NASA Jet Propulsion Laboratory)
Siv K. Lauvset (NORCE Research / Bjerknes Centre for Climate Research)
Tim DeVries (University of California, Santa Barbara)
Judith Hauck (Alfred Wegener Institute, Helmholtz Centre for Polar and Marine Research)
Christian Rödenbeck (Max Planck Institute for Biogeochemistry)
Nicolas Metzl (Sorbonne Université / LOCEAN)
Andrea J. Fassbender (NOAA Pacific Marine Environmental Laboratory)
Jean-Pierre Gattuso (Sorbonne Université / CNRS; Laboratoire d’Océanographie de Villefranche)
Peter Landschützer (Max Planck Institute for Meteorology)
Rik Wanninkhof (NOAA Atlantic Oceanographic and Meteorological Laboratory)
Christopher Sabine (University of Hawaii at Mānoa)
Simone R. Alin (NOAA Pacific Marine Environmental Laboratory)
Mario Hoppema (Alfred Wegener Institute)
Are Olsen (University of Bergen / Bjerknes Centre for Climate Research)
Matthew P. Humphreys (University of East Anglia)
Kunal Chakraborty (National Institute of Oceanography, India)
Ana C. Franco (University of Miami)
Kumiko Azetsu-Scott (Bedford Institute of Oceanography / Fisheries and Oceans Canada)
Dorothee C. E. Bakker (University of East Anglia)
Leticia Barbero (NOAA Atlantic Oceanographic and Meteorological Laboratory)
Nicholas R. Bates (Bermuda Institute of Ocean Sciences / Arizona State University)
Nicole Besemer (University of Natural Resources and Life Sciences Vienna)
Henry C. Bittig (GEOMAR Helmholtz Centre for Ocean Research Kiel)
Albert E. Boyd (University of Tasmania)
Daniel Broullón (Spanish Institute of Oceanography, IEO-CSIC)
Wei-Jun Cai (University of Delaware)
Brendan R. Carter (University of Washington)
Thi-Tuyet-Trang Chau (LSCE, CEA-CNRS-UVSQ)
Chen-Tung Arthur Chen (National Sun Yat-sen University)
Frédéric Cyr (Fisheries and Oceans Canada)
John E. Dore (University of Hawaii)
Ian Enochs (NOAA Atlantic Oceanographic and Meteorological Laboratory)
Richard A. Feely (NOAA Pacific Marine Environmental Laboratory)
Hernan E. Garcia (NOAA National Centers for Environmental Information)
Marion Gehlen (LSCE, CEA-CNRS-UVSQ)
Prasanna Kanti Ghoshal (CSIR-National Institute of Oceanography, India)
Lucas Gloege (Princeton University)
Melchor González-Dávila (University of Las Palmas de Gran Canaria)
Nicolas Gruber (ETH Zürich)
Debby Ianson (Fisheries and Oceans Canada / Institute of Ocean Sciences)
Yosuke Iida (Japan Meteorological Agency)
Masao Ishii (Meteorological Research Institute, Japan)
Apurva Padamnabh Joshi (CSIR-National Institute of Oceanography, India)
Esther Kennedy (NOAA Pacific Marine Environmental Laboratory)
Alex Kozyr (NOAA National Centers for Environmental Information)
Nico Lange (GEOMAR Helmholtz Centre for Ocean Research Kiel)
Claire Lo Monaco (Sorbonne Université / LOCEAN)
Derek P. Manzello (NOAA Atlantic Oceanographic and Meteorological Laboratory)
Galen A. McKinley (Columbia University / Lamont-Doherty Earth Observatory)
Natalie M. Monacci (NOAA Pacific Marine Environmental Laboratory)
Xosé A. Padin (Spanish Institute of Oceanography, IEO-CSIC)
Ana M. Palacio-Castro (Instituto de Investigaciones Marinas, CSIC)
Fiz F. Pérez (Spanish Institute of Oceanography, IEO-CSIC)
J. Magdalena Santana-Casiano (University of Las Palmas de Gran Canaria)
Jonathan Sharp (University of Delaware)
Adrienne Sutton (NOAA Pacific Marine Environmental Laboratory)
Jim Swift (Scripps Institution of Oceanography)
Toste Tanhua (GEOMAR Helmholtz Centre for Ocean Research Kiel)
Maciej Telszewski (International Ocean Carbon Coordination Project, IOCCP)
Jens Terhaar (University of Bern)
Ruben van Hooidonk (University of Miami / NOAA Coral Reef Watch)
Anton Velo (Spanish Institute of Oceanography, IEO-CSIC)
Andrew J. Watson (University of Exeter)
Angelicque E. White (Oregon State University)
Zelun Wu (University of Delaware)
Liang Xue (Xiamen University)
Hyelim Yoo (University of Maryland / NOAA NCEI)
Jiye Zeng (National Institute for Environmental Studies, Japan)
Guorong Zhong (Xiamen University)