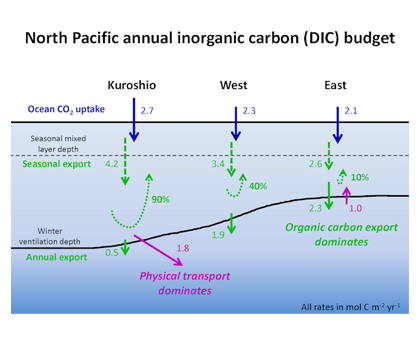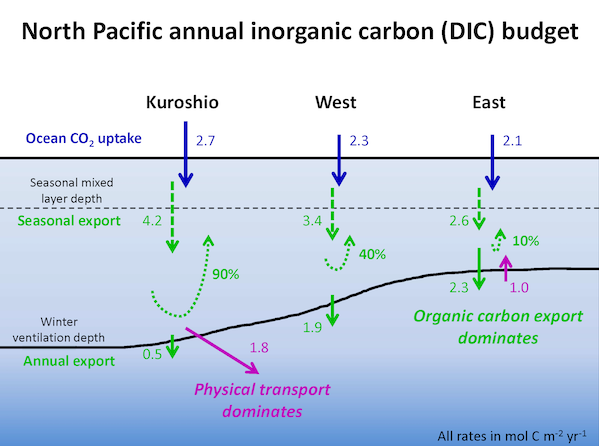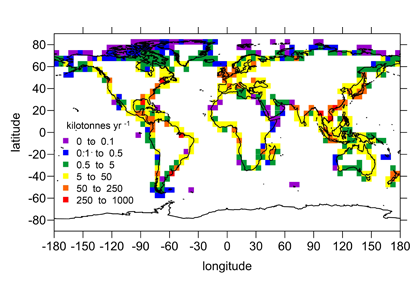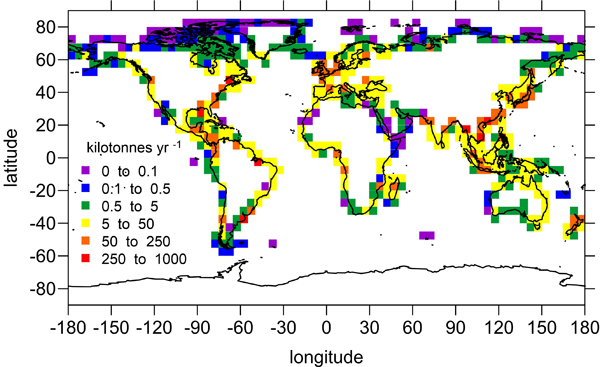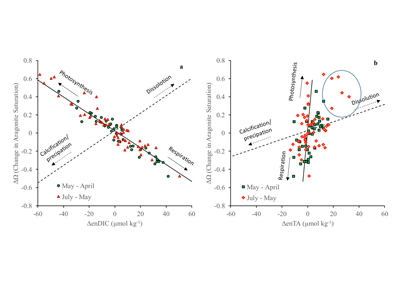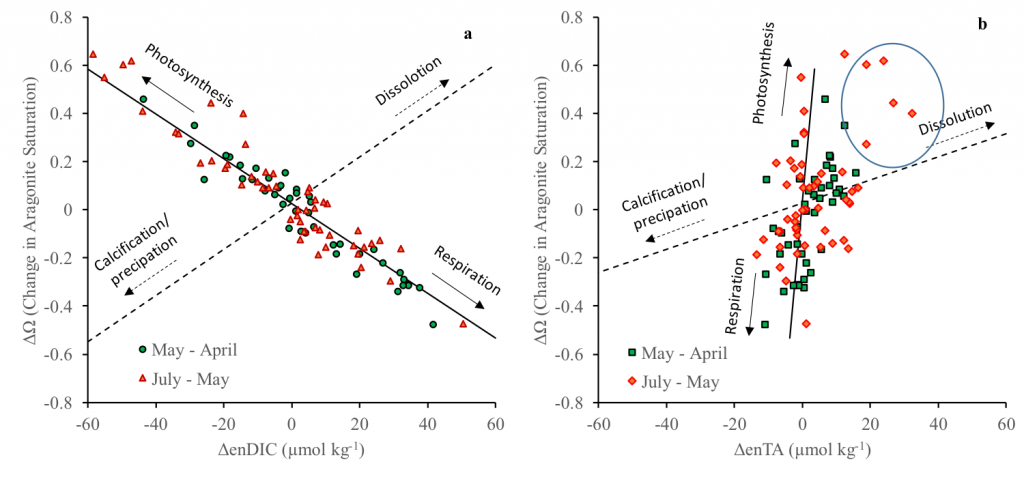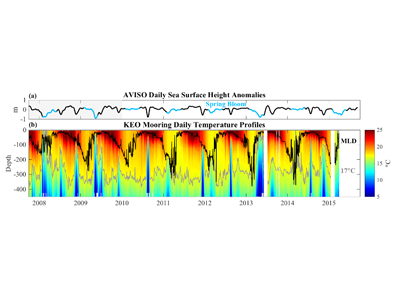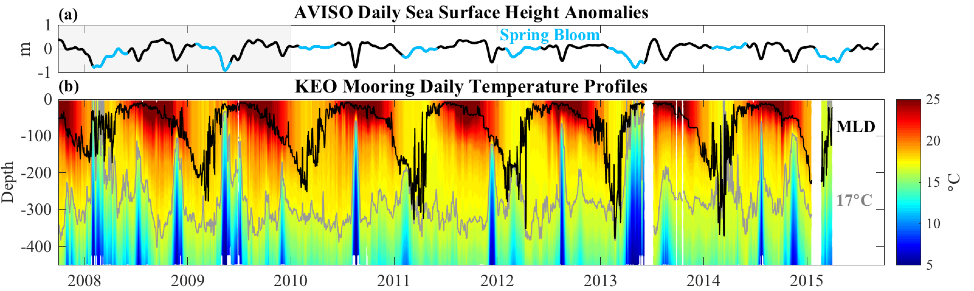
Under the increasing threat of climate change, conservation practitioners and policy makers are seeking innovative and data–driven recommendations for mitigating emissions and increasing natural carbon sinks through nature-based solutions. While the ocean and terrestrial forests, and more recently, coastal wetlands, are well known carbon sinks, there is interest in exploring the carbon storage potential of other coastal and marine ecosystems such as coral reefs, kelp forests, phytoplankton, planktonic calcifiers, krill, and teleost fish. A recent study in Frontiers in Ecology and the Environment reviewed the potential and feasibility of managing these other coastal and marine ecosystems for climate mitigation. The authors concluded, that while important parts of the carbon cycle, coral reefs, kelp forests, planktonic calcifiers, krill, and teleost fish do not represent long-term carbon stores, and in the case of fish, do not represent a sequestration pathway. Phytoplankton do sequester globally significant amounts of carbon and contribute to long-term carbon storage in the deep ocean, but there is currently no good way to manage them to increase their carbon storage capacity; additionally, the vast majority of phytoplankton is located in international waters that are outside national jurisdictions, making it very difficult to include them in current climate mitigation policy frameworks.
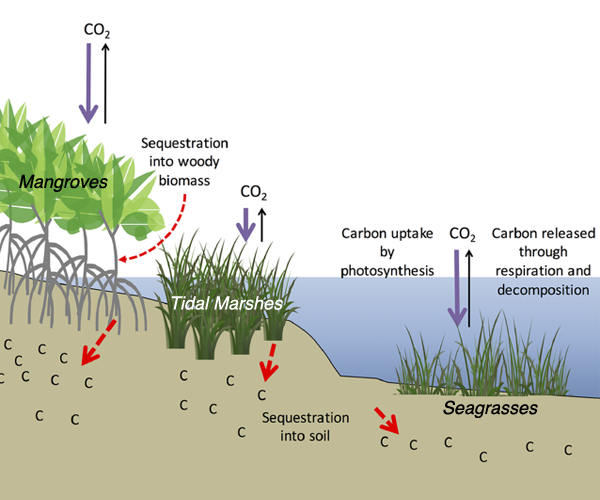
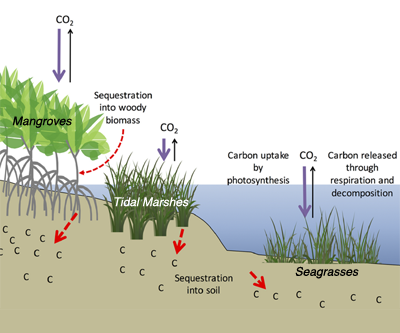
Comparatively, coastal wetlands (mangroves, tidal marshes, and seagrasses) effectively sequester carbon long-term (up to 10x more carbon stored per unit area than terrestrial forests with 50-90% of the stored carbon residing in the soil), and fall within clear national jurisdictions, which facilitates effective and quantifiable management actions. In addition, wetland degradation has the potential to release vast amounts of stored carbon back into the atmosphere and water column, meaning that conservation and restoration of these systems can also reduce potential emissions. The authors conclude that coastal wetland protection and restoration should be a primary focus in comprehensive climate change mitigation plans along with reducing emissions.
Authors:
Jennifer Howard (Conservation International)
Ariana Sutton-Grier (University of Maryland, NOAA)
Dorothée Herr (IUCN)
Joan Kleypas (NCAR)
Emily Landis (The Nature Conservancy)
Elizabeth Mcleod (The Nature Conservancy)
Emily Pidgeon (Conservation International)
Stefanie Simpson (Restore America’s Estuaries)
Original paper: http://onlinelibrary.wiley.com/doi/10.1002/fee.1451/full