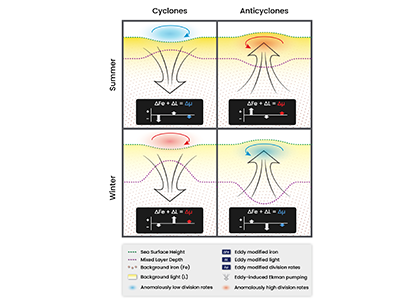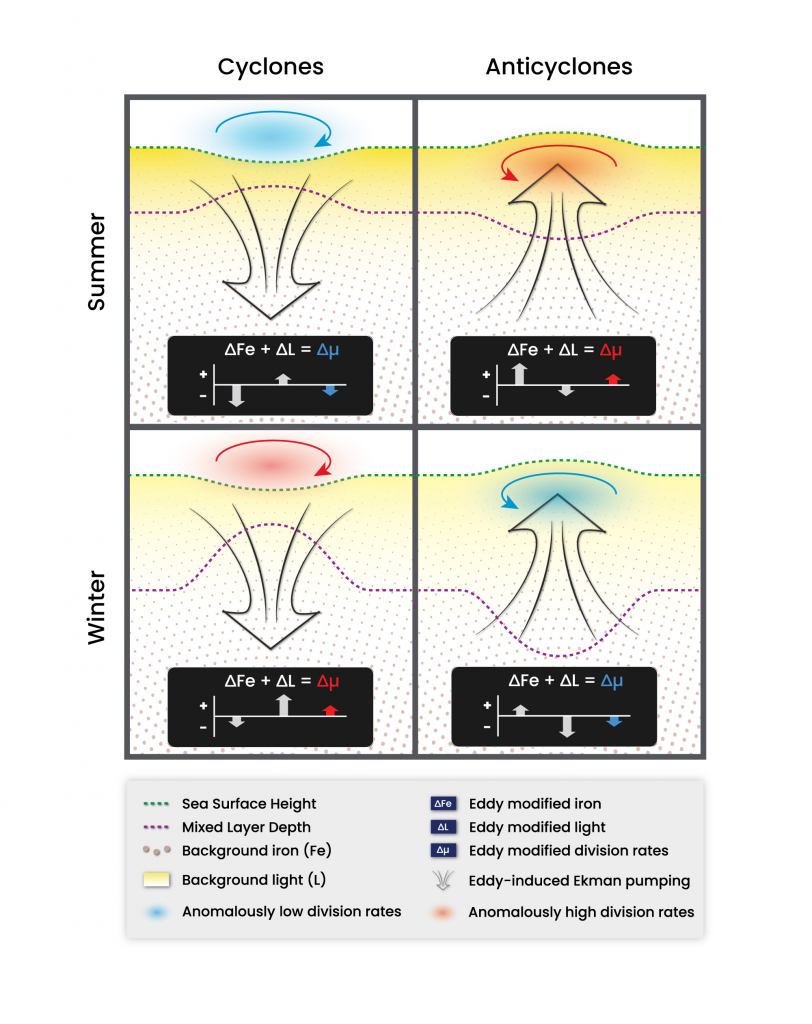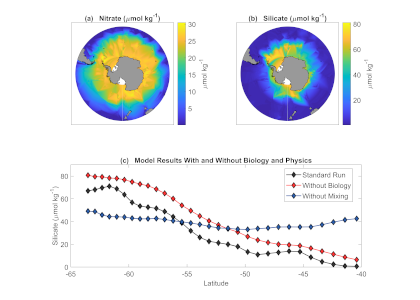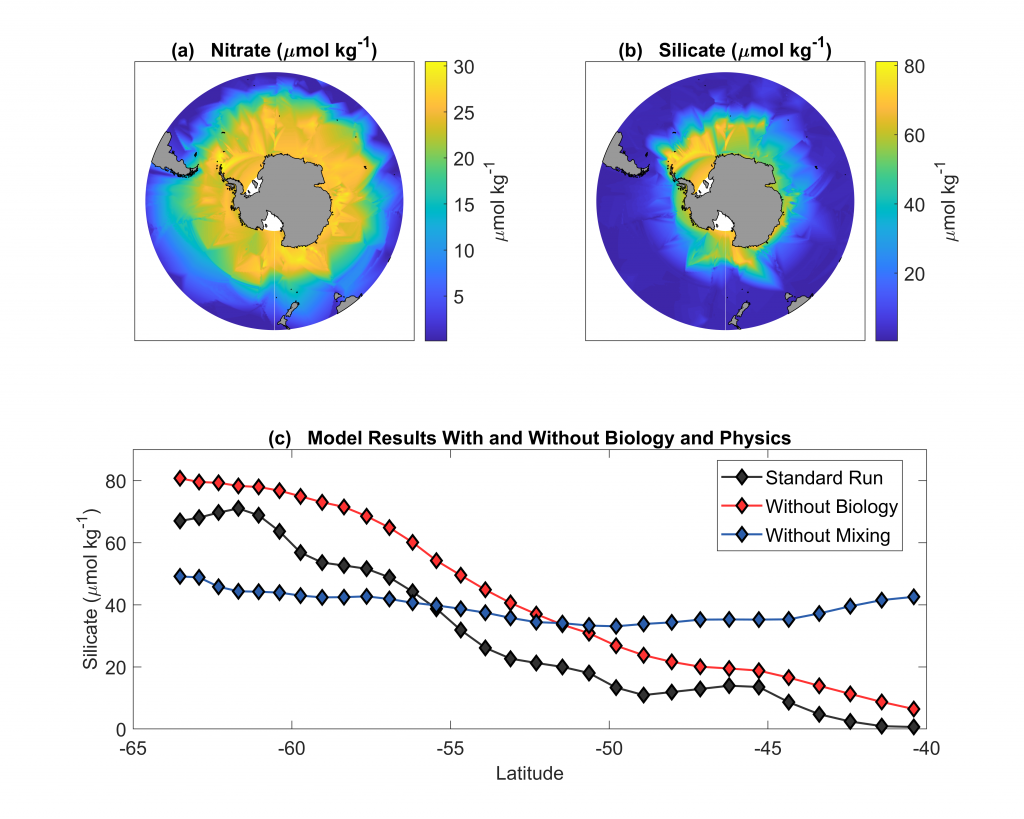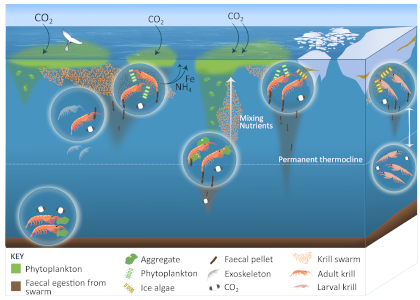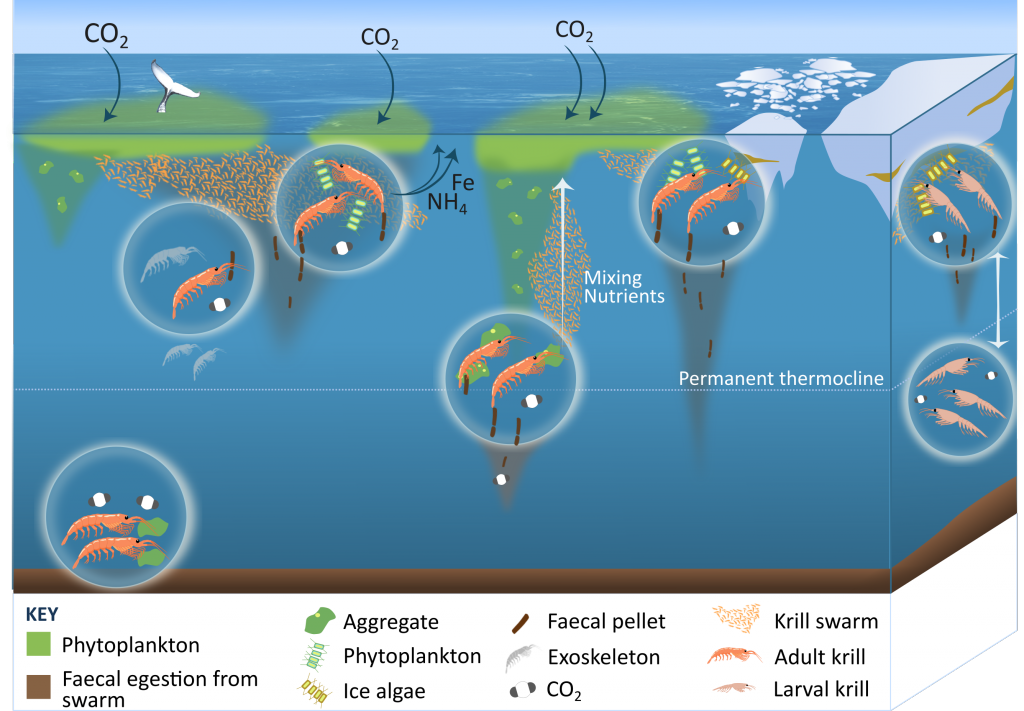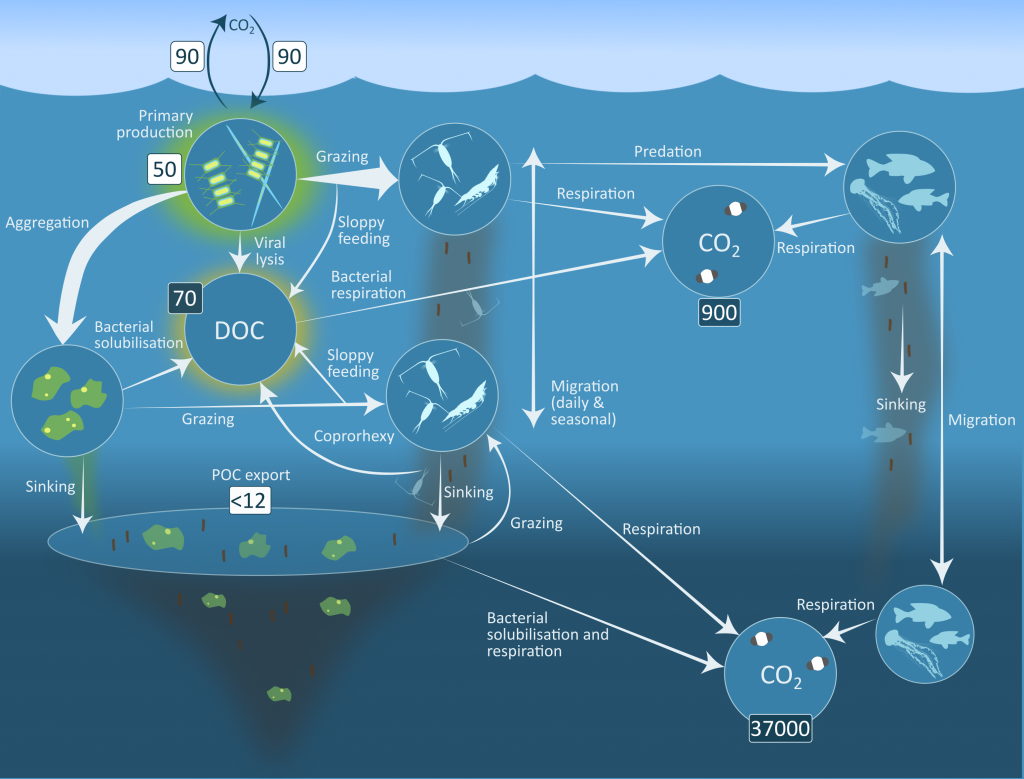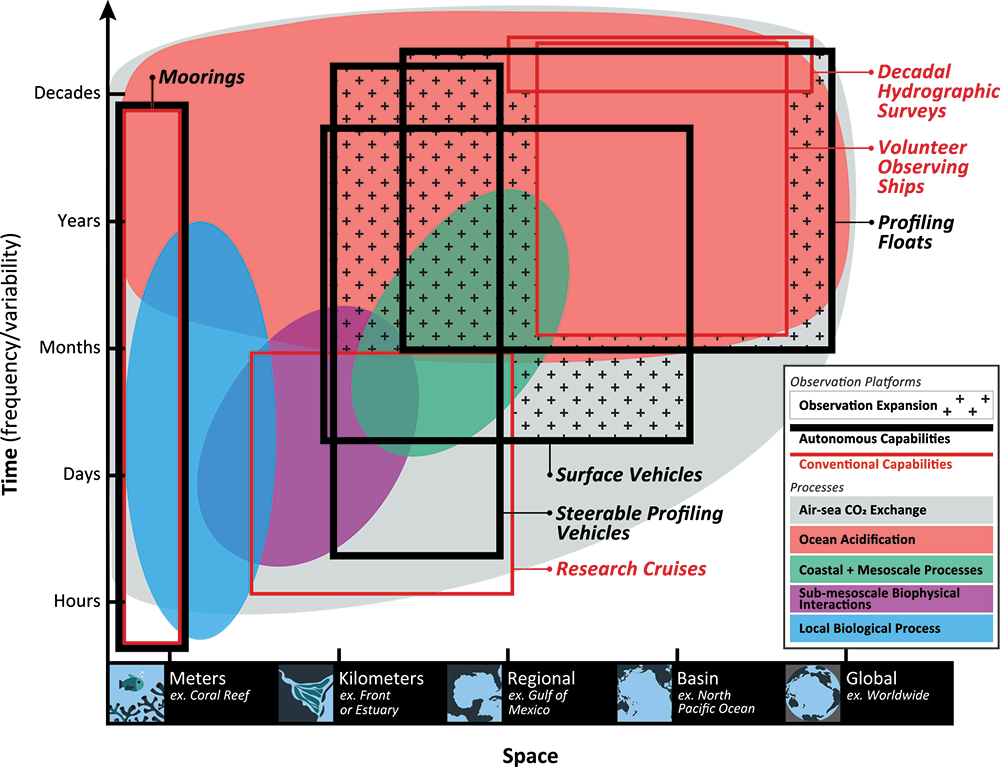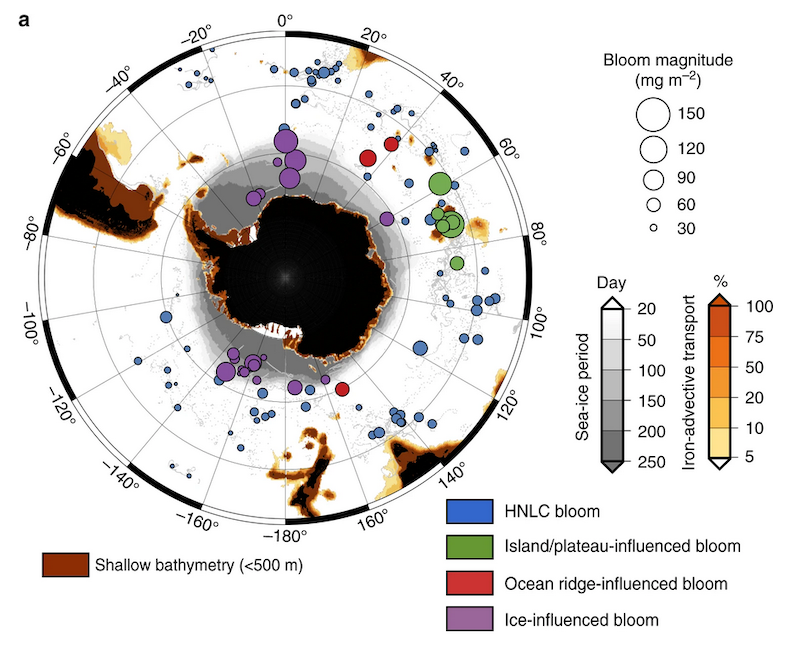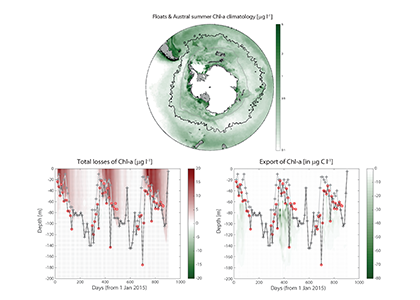
More observations are needed to constrain the relative roles of physical (advection), biogeochemical (downward export), and ecological (grazing and biological losses) processes in driving the fate of phytoplankton blooms in Southern Ocean waters. In a recent paper published in Nature Communications, authors used seven Biogeochemical Argo (BGC-Argo) floats that vertically profiled the upper ocean every ten days as they drifted for three years across the remote Sea Ice Zone of the Southern Ocean. Using the floats’ biogeochemical sensors (chlorophyll, nitrate, and backscattering) and regional ratios of nitrate consumption:chlorophyll synthesis, the authors developed a new approach to remotely estimate the fate of the phytoplankton stocks, enabling calculations of herbivory and of downward carbon export. The study revealed that the major fate of phytoplankton biomass in this region is grazing, which consumes ~90% of stocks. The remaining 10% is exported to depth. This pattern was consistent throughout the entire sea ice zone where the floats drifted, from 60°-69° South.
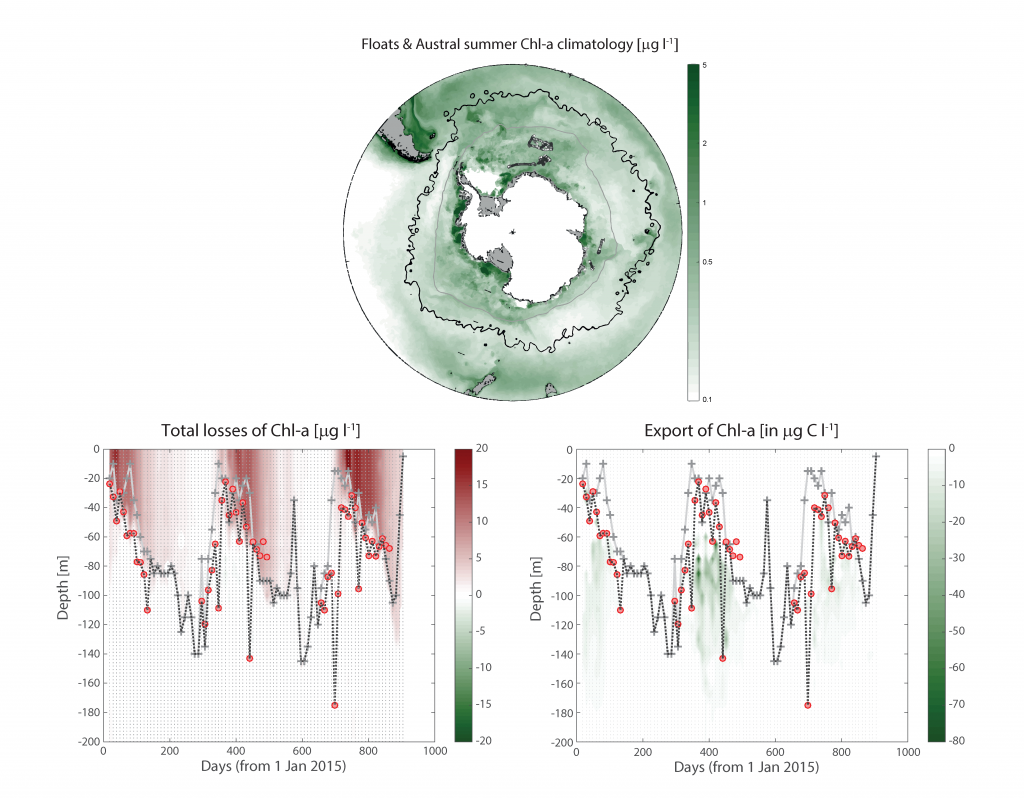
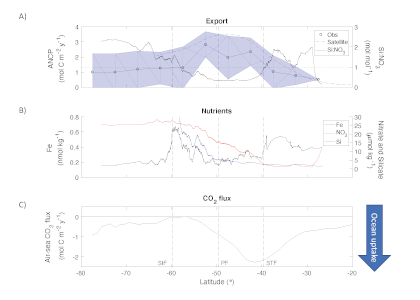
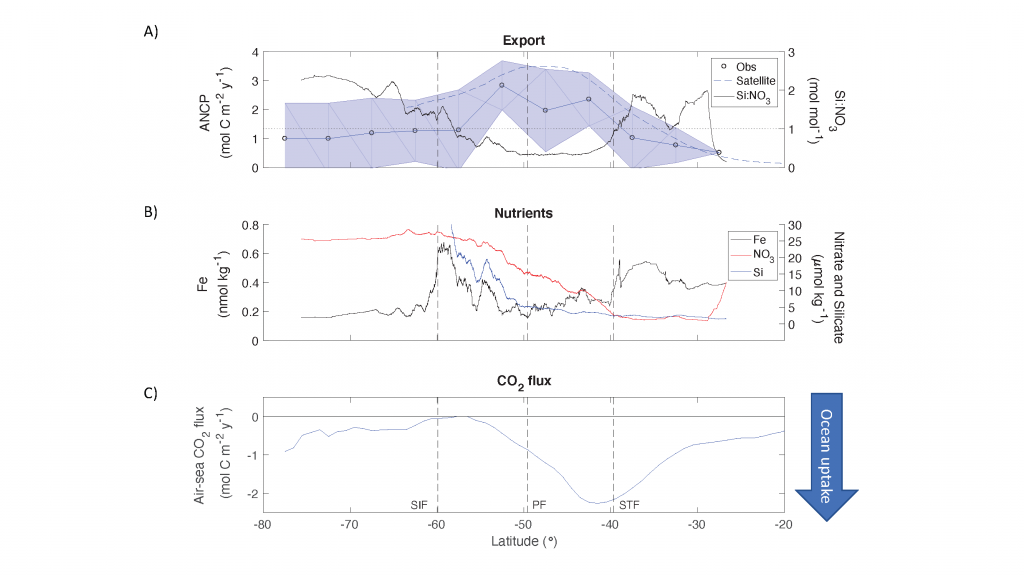
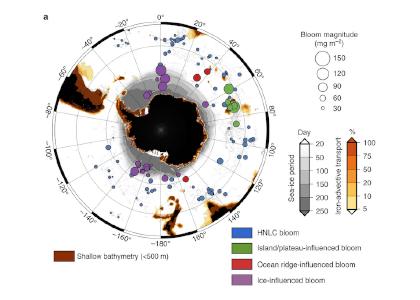
Figure Caption: Southern Ocean Chlorophyll a climatology and floats’ trajectories (top panel). Total losses of Chlorophyll a (including grazing and phytodetritus export, left panel). Phytodetritus export (right panel).
This study region comprises two of the three major krill growth and development areas—the eastern Weddell and King Haakon VII Seas and Prydz Bay and the Kerguelen Plateau—so the observed grazing was probably due to Antarctic krill, underscoring their pivotal importance in this ecosystem. Building upon the greater understanding of ocean ecosystems via satellite ocean colour development in the 1990s, BGC-Argo floats and this new approach will allow remote monitoring of the different fates of phytoplankton stocks and insights into the status of the ecosystem.
Authors:
Sebastien Moreau (Norwegian Polar Institute, Tromsø, Norway)
Philip Boyd (Institute for Marine and Antarctic Studies, Hobart, Australia)
Peter Strutton (Institute for Marine and Antarctic Studies, Hobart, Australia)