Blue holes are unique depositional environments that are formed within carbonate platforms. Due to an enclosed geomorphology that restricts water exchange, blue hole ecosystems are typically characterized by steep biogeochemical gradients and distinctive microbial communities. For the past three decades, studies have described vertical gradients in physical, chemical, and biological parameters that typify blue hole water columns, but their elemental cycles, particularly carbon, remain poorly understood.

Figure 1. Aerial photo of the Yongle Blue Hole in the South China Sea (Credit: P. Yao et al./JGR Biogeosciences)
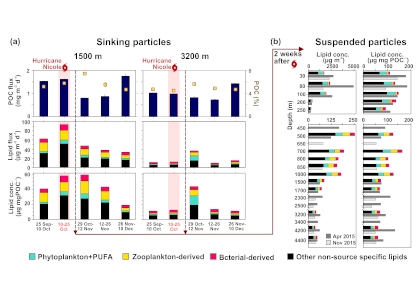
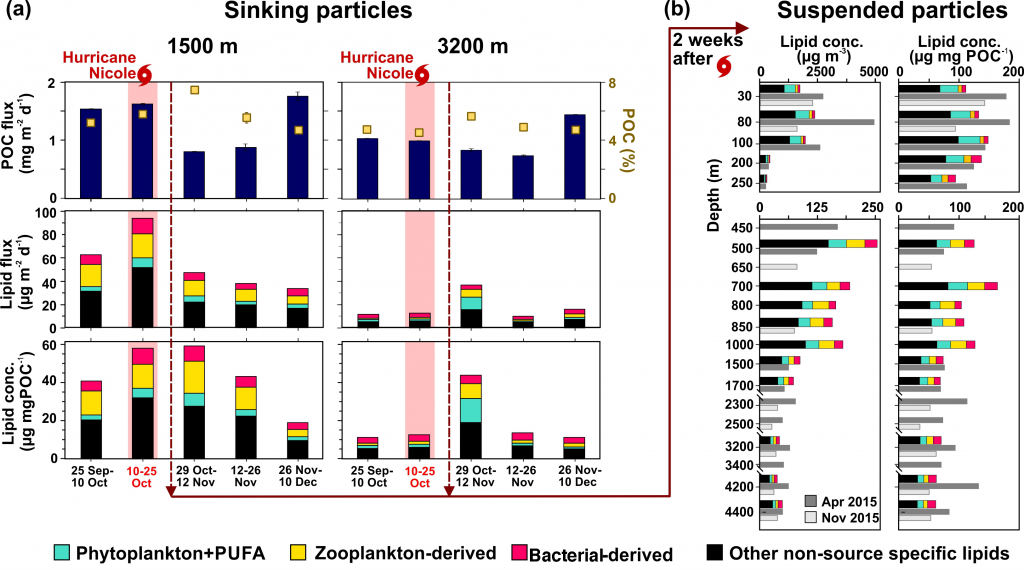
In July 2016, the Yongle Blue Hole (YBH) was discovered to be the deepest known blue hole on Earth (~300 m). YBH is located in the Xisha Islands of the South China Sea. The unique features and ease of accessibility make YBH an ideal natural laboratory for studying carbon cycling in marine anoxic systems. In a recent study published in JGR Biogeosciences, the authors reported extremely low concentrations of dissolved organic carbon (DOC) (e.g., 22 µM) and very high concentrations of dissolved inorganic carbon (DIC) (e.g., 3,090 µM) in YBH deep waters. Radiocarbon dating revealed that the YBH DOC and DIC were unusually old, yielding ages (6,810 and 8270 years BP, respectively) that are much more typical of open ocean deep water. Based on H2S and microbial community composition profiles, the authors concluded that sharp redox gradients and a high abundance of sulfur cycling bacteria were likely responsible for much of the DOC consumption in YBH. The unusually low concentrations and old DOC ages in the relatively shallow YBH suggest short-term cycling of recalcitrant DOC in oceanic waters, which has been recognized as a long-term microbial carbon sink in the global ocean. The stoichiometry of DIC and total alkalinity changes suggested that the accumulation of DIC in the deep layer of the YBH was largely derived from both the dissolution of carbonate and OC decomposition through sulfate reduction. However, the role of carbonate dissolution from the walls of the blue hole in affecting the old ages of carbon in this system remain uncertain, yet there appears to no evidence of subterranean freshwater into the bottom waters of the blue hole. In the face of expanding oxygen minimum zones and anthropogenically-induced coastal hypoxia, blue holes such as YBH can provide an accessible natural laboratory in which to study the microbial and biogeochemical features that typify these low-oxygen systems.
Authors:
Peng Yao (Ocean University of China)
Thomas S. Bianchi (University of Florida)
Xuchen Wang (Ocean University of China)
Zuosheng Yang (Ocean University of China)
Zhigang Yu (Ocean University of China)