The OCB Benthic Ecosystem & Carbon Synthesis (BECS) are hosting a webinar!
WHEN
November 28, 2023, 10:00-11:00 AM EST
SPEAKERS
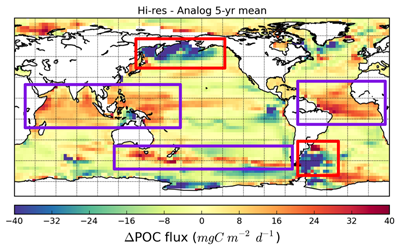
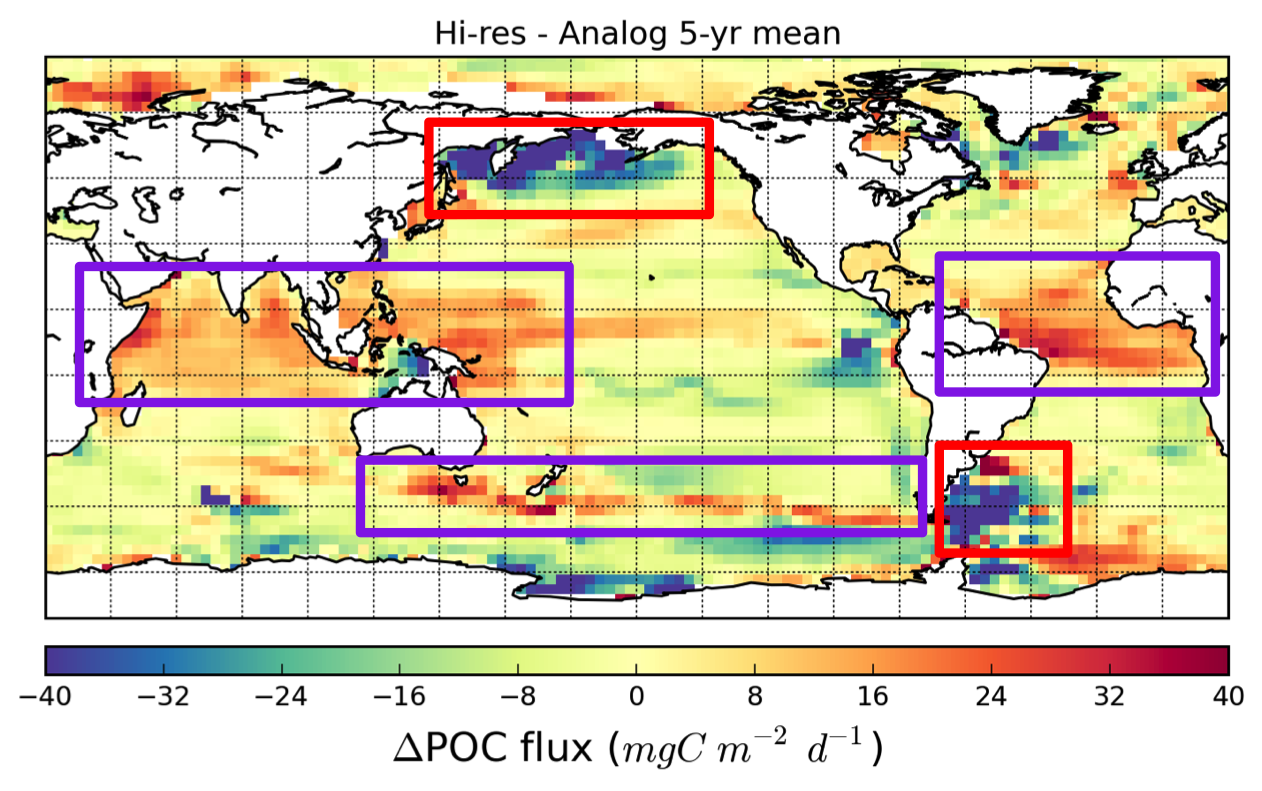
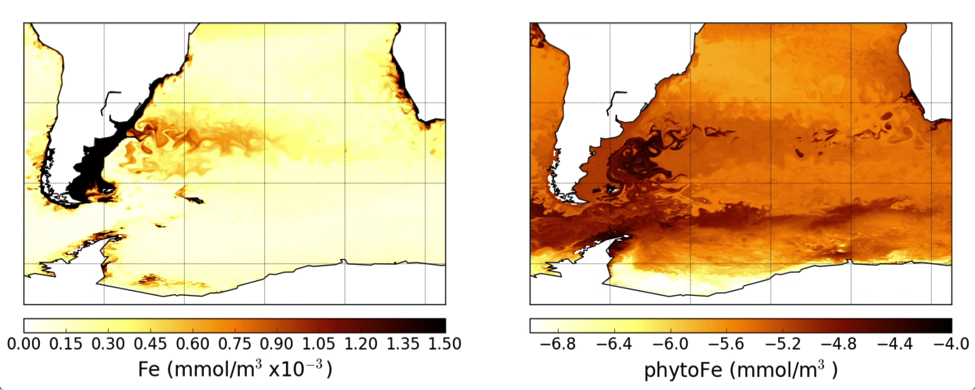
Chris Somes (GEOMAR) - The impact of reductive sedimentary iron release on changing ocean biogeochemistry simulations of the Anthropocene
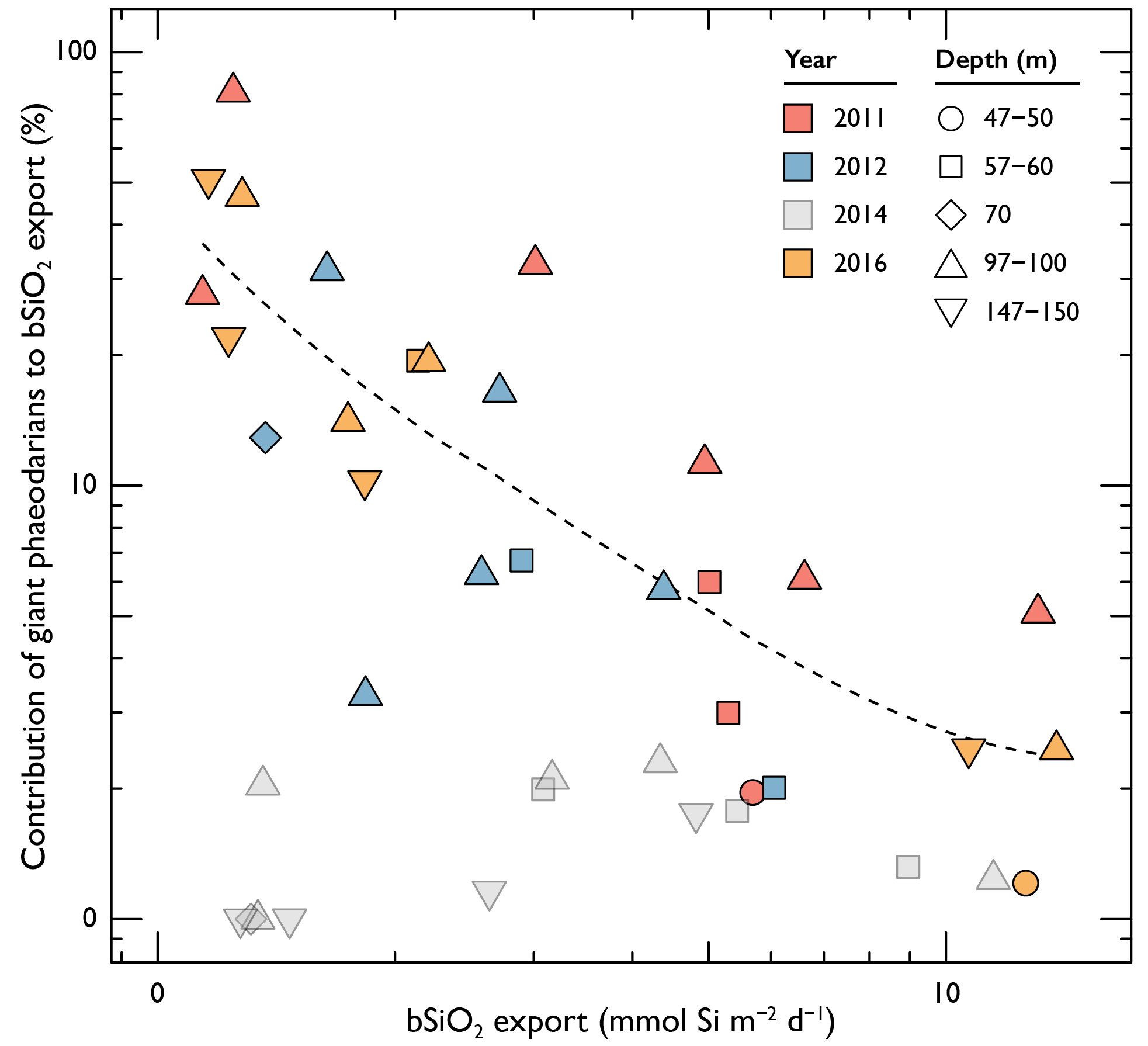
Kanchan Maiti (LSU) - Oxygen and carbon dynamics in Mississippi river influenced shelf sediments