How will increasing meltwater from Greenland affect the biogeochemistry of the ocean? Release of meltwater into the ocean has physical and biogeochemical effects on the local water column. With respect to nutrient availability, meltwater supplies the bioessential nutrients iron and silicic acid but is deficient in nitrate and phosphate. However, despite very low meltwater nitrate and phosphate concentrations, pronounced summertime phytoplankton blooms are observed in many, though not all, of Greenland’s large fjord systems. These unusual summertime blooms are associated with meltwater from marine-terminating glaciers. So if the meltwater itself is not supplying nitrate and phosphate that these blooms require, what is the source of the nutrients that support these blooms?
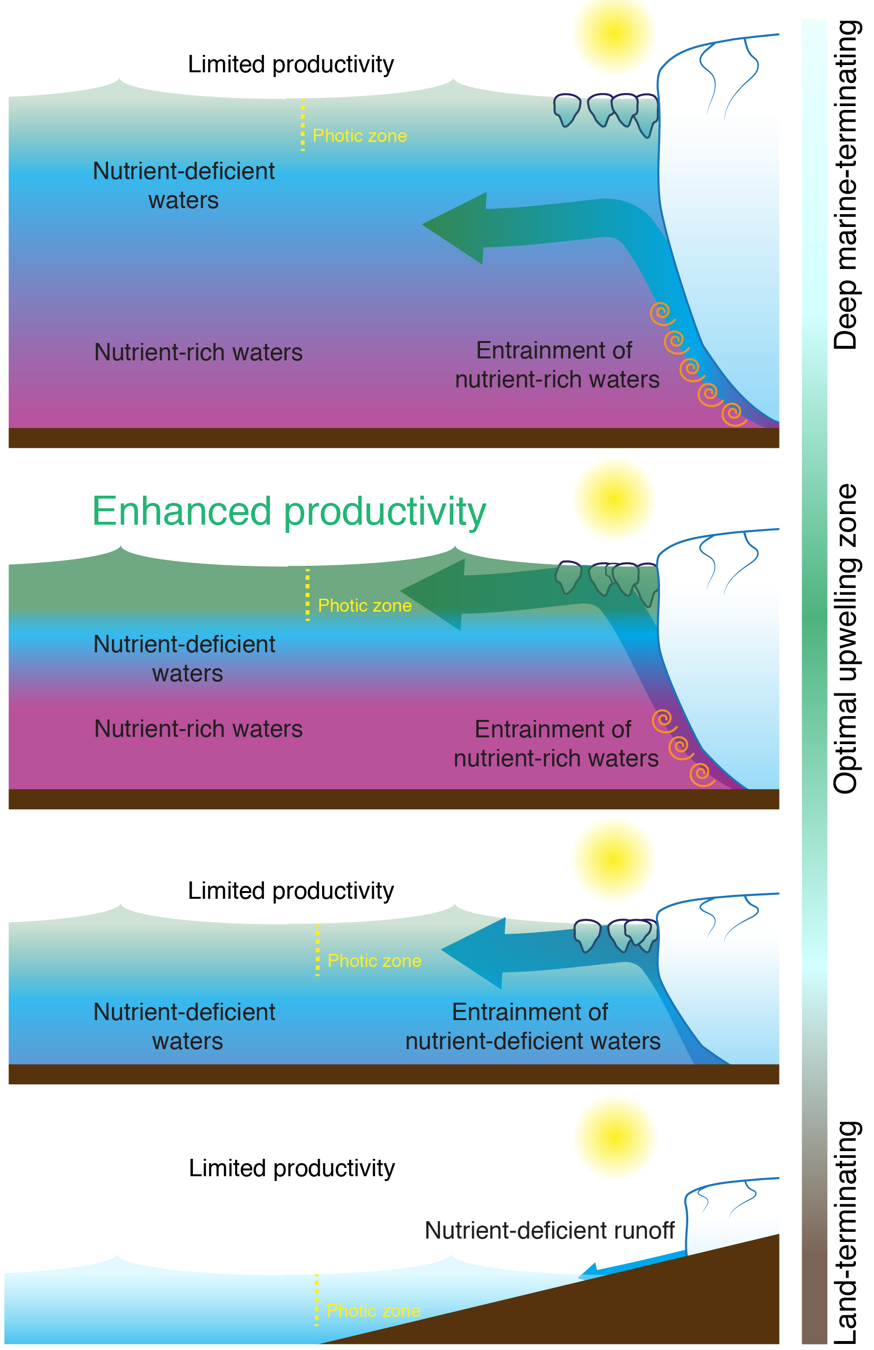
An illustration of how changing the depth of a glacier affects downstream productivity
A recent study published in Nature Communications shows that when meltwater is released below sea level under large marine-terminating glaciers, it rises rapidly towards the surface in buoyant discharge plumes. As these plumes rise, they entrain large quantities of deep, nutrient-rich seawater. This vertical transport, or ‘pumping’, of these nutrients to the surface sustains unusually high summertime productivity in Greenland’s fjords. Conversely, when meltwater is released at the ocean surface, primary production is reduced because the meltwater itself lacks the nitrate and phosphate required to fuel phytoplankton blooms. Consequently, the inland retreat of Greenland’s large marine-terminating glaciers is ultimately bad news for summertime marine phytoplankton communities. As the depth of the marine-terminating glaciers shoals, their associated nutrient ‘pumps’ collapse, which will likely have negative effects on primary production and associated inshore fisheries.
Authors:
M.J. Hopwood (GEOMAR)
D. Carroll (Jet Propulsion Laboratory)
T.J. Browning (GEOMAR)
L. Meire (Royal Netherlands Institute for Sea Research and Greenland Climate Research Centre)
J. Mortensen (Greenland Climate Research Centre)
S. Krisch (GEOMAR)
E.P. Achterberg (GEOMAR)