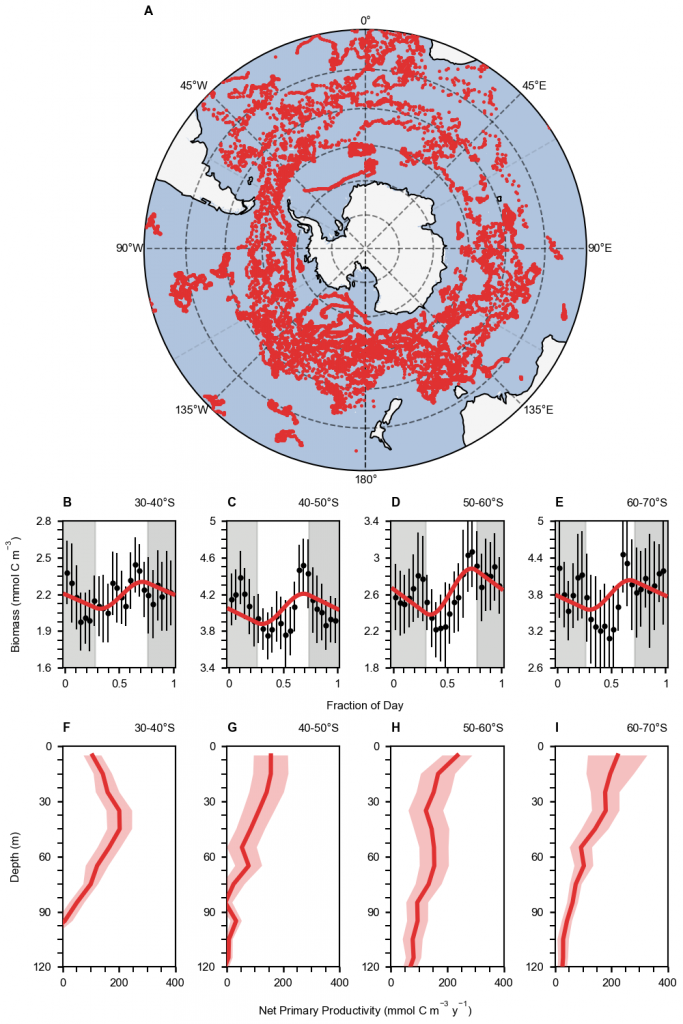
The Deepwater Horizon oil spill (April-July 2010) in the NE Gulf of Mexico provided researchers with an opportunity to explore what happens when marine snow and oil mix. Marine snow are detrital particles or aggregates consisting of inorganic and organic components, such as bacteria, phytoplankton cells, zooplankton fecal pellets, and mucous feeding webs, and are important in the biological pump and export of carbon to deep water. It is now known that marine snow and oil interact to form marine-oil-snow (MOS) which sediments to the seafloor, supported by observations from experiments, sediment traps, and sediment cores.
In a recent study published in the Journal of Geophysical Research Oceans, the authors provide additional analyses of the impact of oil on marine snow. The SIPPER camera imaging system was deployed on 13 cruises between May 2010 and August 2014 (during and after the oil spill), collecting more than 117 million images of aggregates. Analyses of these images indicated that diatom chains and Acantharian (small animals) spines were relatively common components of aggregates. The oil spill, combined with high Mississippi River outflow, resulted in marine snow concentrations that were significantly higher with larger-sized particles during the oil spill than in follow-on years. The shape of particles was consistently elongated in all years compared to the spherical shape assumed for simulations of particle sinking speeds. Analysis of the fractal dimension or surface roughness of particles indicated that during the oil spill (May 2010) aggregates had significantly higher fractal dimensions, suggesting that oil droplets in the marine-oil-snow reduced the amount of empty space within aggregates, thereby increasing particle density and increasing the sedimentation of oil. Fractal dimensions also increased with particle size in all years and, therefore, was not an impact of the oil spill. These data provide a baseline for future biogeochemical studies in the northern Gulf of Mexico and for model development for future oil spill response scenarios.
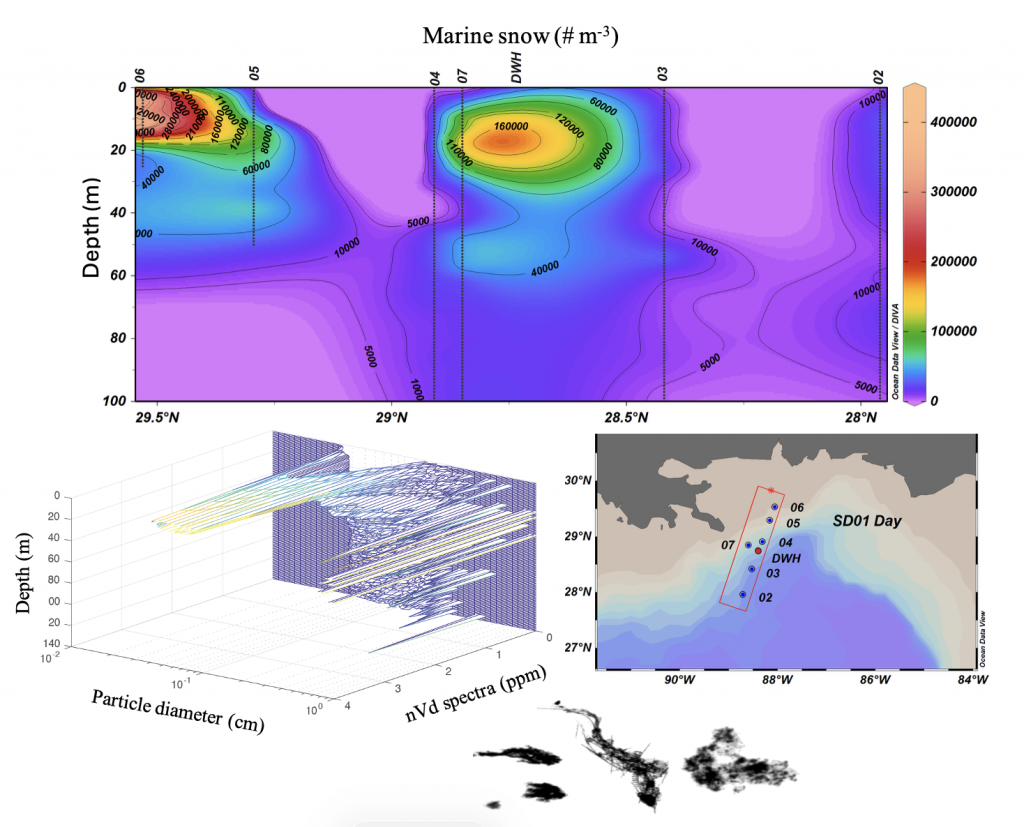
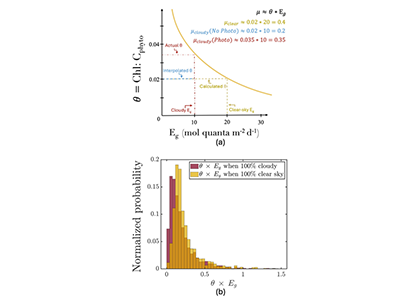
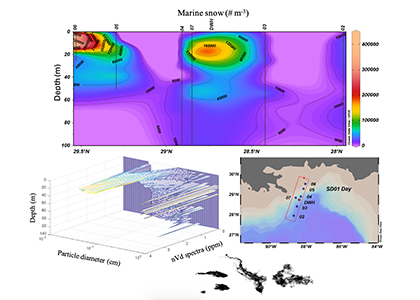
Figure caption. The abundance and distribution of marine snow was spatially variable, but unusually high in the upper 20 m of the water column during the summer following the Deepwater Horizon (DWH) oil spill (upper panel). Previously reported concentrations were 1,000 – 6,000 particles m-3. High concentrations occurred at the DWH platform site and shelf edge stations (lower right panel). Smaller sized particles were abundant near surface with larger particles (up to 1 cm) observed deeper in the water column (lower left panel). Examples of marine snow images are shown at the bottom. nVd is the normalized particle volume spectra, d is the median diameter within each particle size bin.
Authors:
Kendra L. Daly (University of South Florida)
George Jackson (Texas A&M University)
Andrew Remsen (Bureau of Ocean Energy Management)
Kurt Kramer (OceanSpace Sensors)
Palak Dave (Moffitt Cancer Center and Research Institute)
Dmitry B. Goldgof (University of South Florida)
Lawrence Hall (University of South Florida)
Citation:
Daly, K. L., Jackson, G., Remsen, A., Kramer, K., Dave, P., Goldgof, D. B., & Hall, L. (2026). Marine snow dynamics in the NE Gulf of Mexico: Particle abundance, characteristics, and impacts on Deepwater Horizon oil sedimentation. Journal of Geophysical Research: Oceans, 131, e2025JC023316. https://doi.org/10.1029/2025JC023316