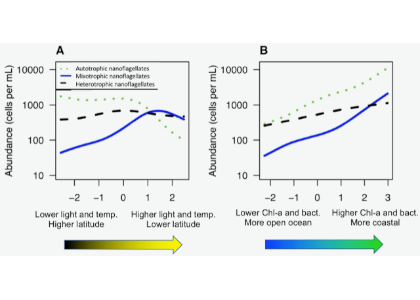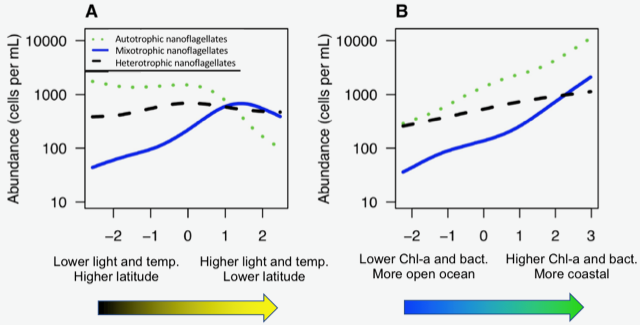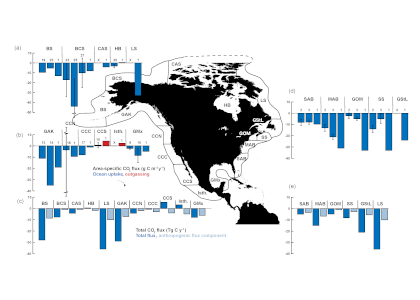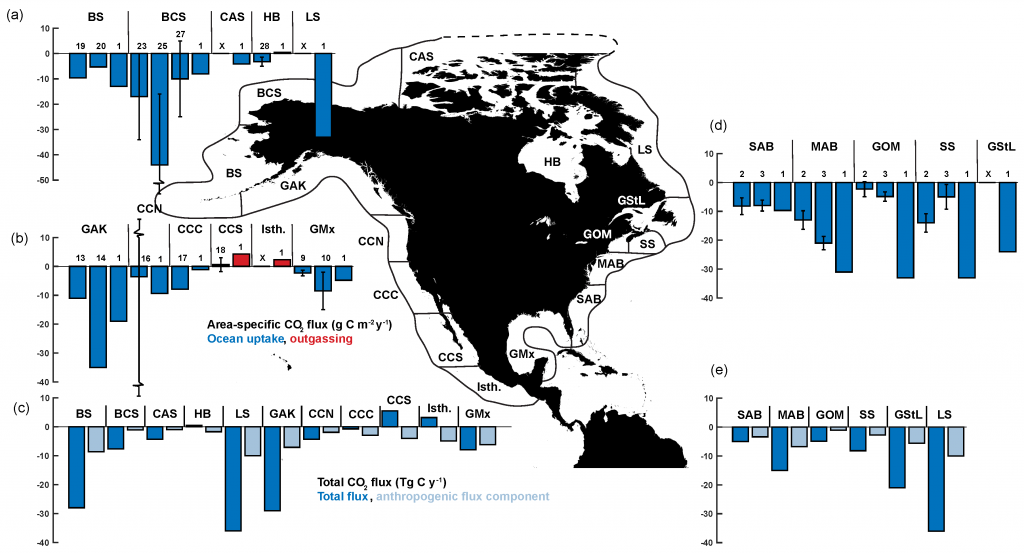
Joint science highlight with GEOTRACES
Carbon storage in the ocean is sensitive to the depths at which particulate organic carbon (POC) is respired back to CO2 within the twilight zone (100-1000m). For decades, it has been an oceanographic priority to determine the depth scale of this regeneration process. To investigate this, GEOTRACES scientists are deploying new isotopic tools that provide a high-resolution snapshot of POC flux and regeneration across steep biogeochemical gradients in the South Pacific Ocean.
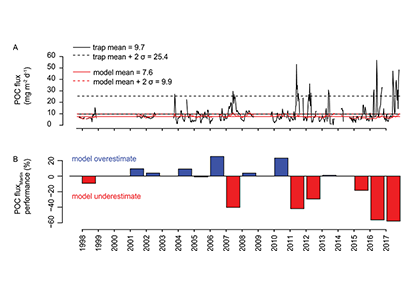
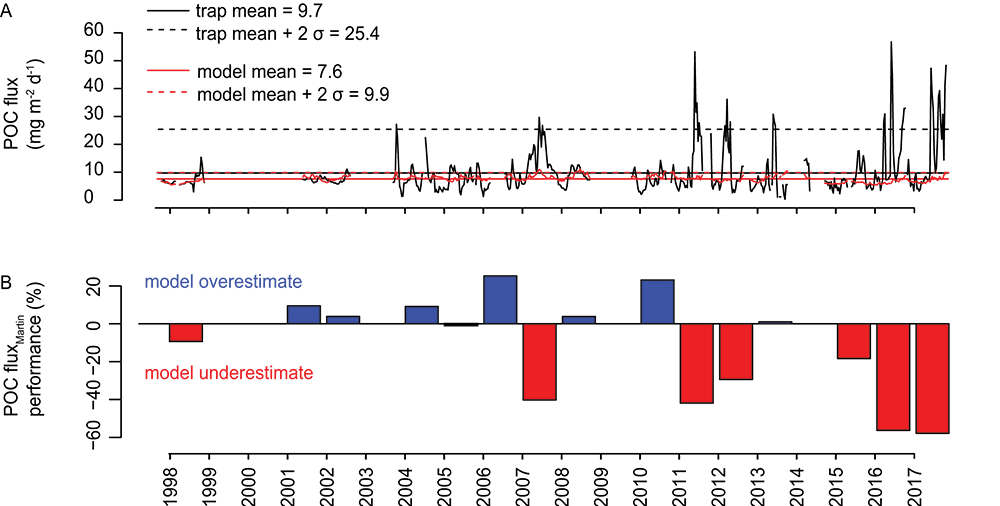
A recent paper in PNAS reported on particulate organic carbon (POC) fluxes throughout the water column (focusing on the upper 1000 m) along the GP16 GEOTRACES section between Peru and Tahiti (Figure 1A). POC fluxes (Figure 1B) were derived by normalizing concentrations of POC to 230Th following analysis of samples collected by in situ filtration. This work builds on a research theme initiated at the GEOTRACES-OCB synthesis workshop held at Lamont-Doherty Earth Observatory in 2016.
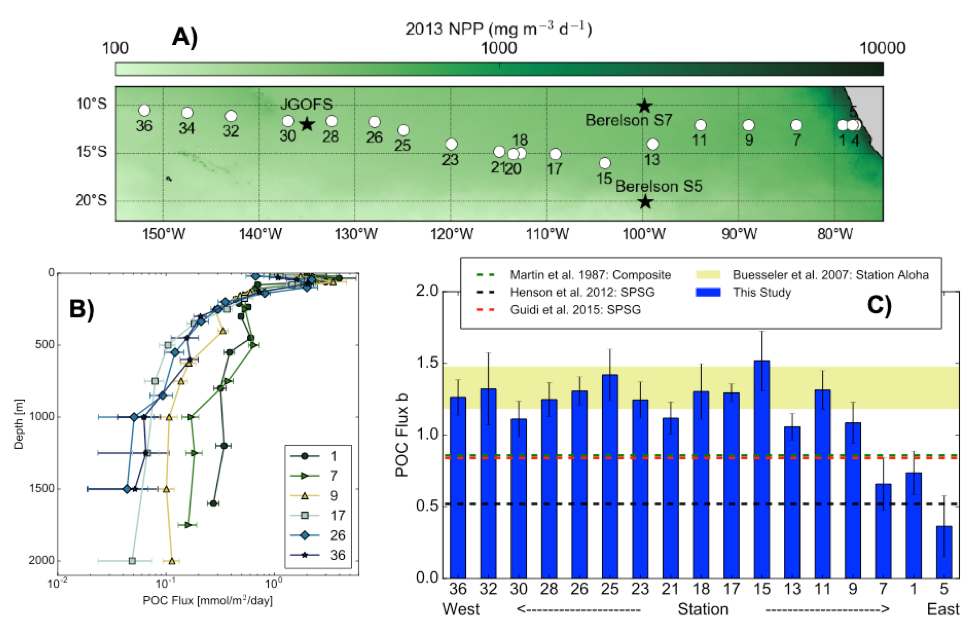
Figure caption: Site map and POC flux characteristics from GEOTRACES GP16 section. Plot A) shows the GP16 station locations as white circles, with nearby sediment trap deployments as black stars, with 2013 MODIS satellite-derived net primary productivity in the background. Plot B) shows POC fluxes from particulate 230Th-normalization from selected stations spanning the zonal extent of the GP16 section. Plot C) shows power law exponent b values for each GP16 station (blue), compared to estimates from bottom-moored sediment traps in the South Pacific (black and red dashed lines), a compilation of sediment traps in the North Pacific (green dashed line), and neutrally buoyant sediment traps in the subtropical North Pacific (yellow shaded band). GP16 regeneration length scales from 230Th-normalization agree most closely with the estimates from neutrally buoyant sediment traps.
The study results show that POC regeneration depth is shallower than anticipated, especially in warm stratified waters of the subtropical gyre. Regeneration depth—expressed in terms of the Martin-curve power-law exponent “b” (Figure 1C)—is shown to be greater than previous estimates (horizontal dashed lines), but similar to values obtained using neutrally buoyant sediment traps at the Hawaii Ocean Time-series Station Aloha. In contrast to the rapid regeneration of POC in warm stratified waters, POC regeneration within the ODZ is below our detection limits. Models have shown that shallower regeneration of POC leads to less efficient carbon storage in the ocean, making the authors speculate that global warming, yielding expanded and more stratified gyres, may induce a reduction of the ocean’s efficacy for carbon storage via the biological pump.
Authors:
Frank J. Pavia, Robert F. Anderson, Sebastian M. Vivancos, Martin Q. Fleisher (Columbia University)
Phoebe J. Lam (University of California Santa Cruz)
B.B. Cael (now at University of Hawai’i Manoa, formerly at MIT)
Yanbin Lu, Pu Zhang, R. Lawrence Edwards (University of Minnesota)
Hai Cheng (University of Minnesota and Xi’an Jiaotong University)