Net primary production (NPP)—the organic carbon produced by the phytoplankton minus the organic carbon respired by phytoplankton themselves—serves as a major energy source of the marine ecosystem. Traditional methods for measuring NPP rely on ship-based discrete sampling and bottle incubations (e.g., 14C incubation), which introduce potential artifacts and limit the spatial and temporal data coverage of the global ocean. The global distribution of NPP has been estimated using satellite observations, but the satellite remote sensing approach cannot provide direct information at depth.
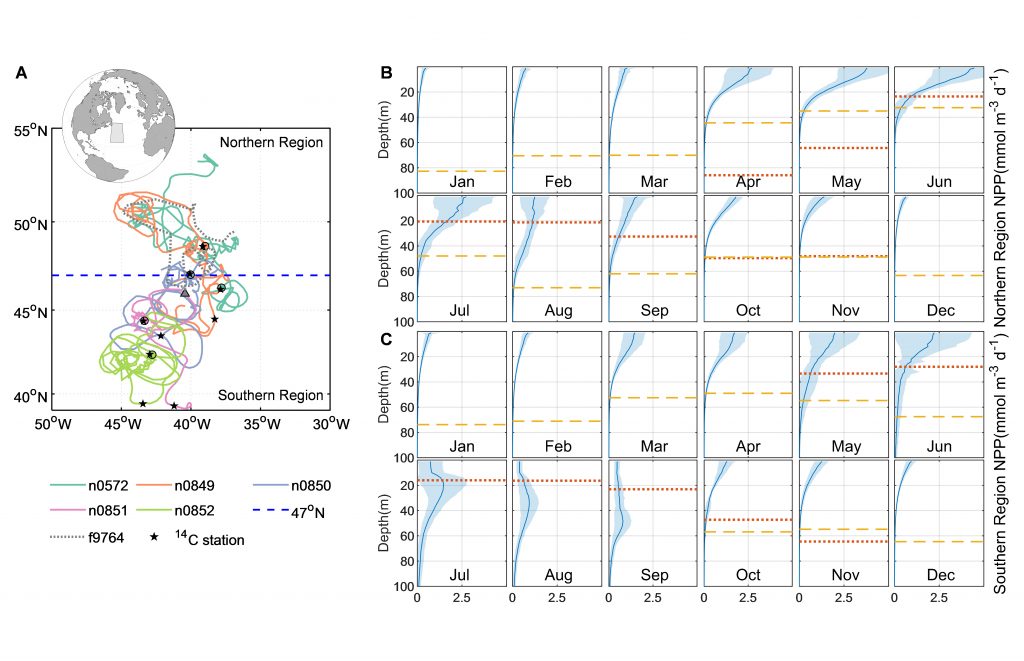
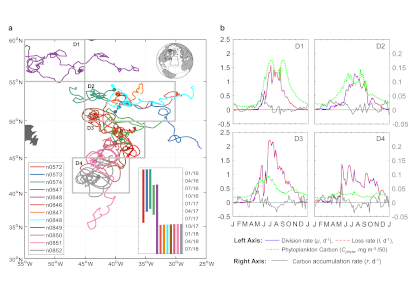
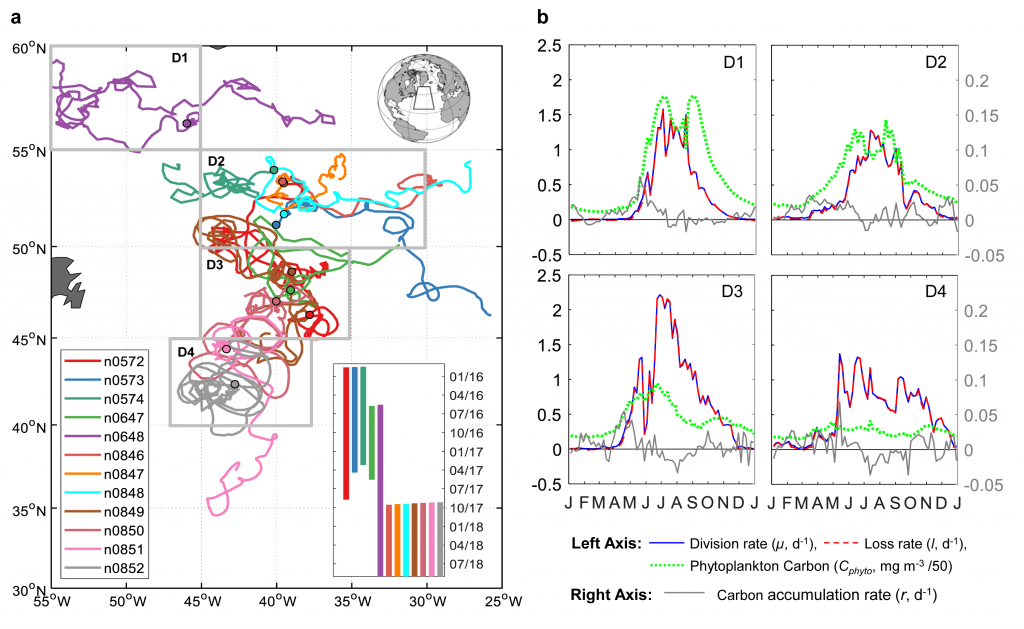
Figure 1. Panel A. Trajectories of 5 BGC-Argo and 1 SOS-Argo with the initial float deployment locations denoted by filled symbols. The dash-line at 47° N divided the research area into the northern (temperate) and southern (subtropical) regions. Stars indicate ship stations where 14C NPP values were measured during NAAMES cruises and compared with NPP from nearby Argo floats. Panels B and C. Monthly climatologies of net primary production (NPP, mmol m-3 d-1) profiles in the northern and southern regions of the research area, derived from BGC-Argo measurements using the PPM model. The shadings indicate one standard deviation. The red dotted line indicates mixed layer depth (MLD, m), and the yellow dashed line shows euphotic depth (Z1%, m).
To fill this niche, a recent study in Journal of Geophysical Research: Biogeosciences, applied bio-optical measurements from Argo profiling floats to study the year-round depth-resolved NPP of the western North Atlantic Ocean (39° N to 54° N). The authors calculated NPP with two bio-optical models (Carbon-based Productivity Model, CbPM; and Photoacclimation Productivity Model, PPM). A comparison with NPP profiles from 14C incubation measurements showed advantages and limitations of both models. CbPM reproduced the magnitude of NPP in most cases, but had artifacts in the summer (a large NPP peak in the subsurface) due to the subsurface chlorophyll maximum caused by photoacclimation. PPM avoided the artifacts in the summer from photoacclimation, but the magnitude of PPM-derived NPP was smaller than the 14C result. Latitudinally varying NPP were observed, including higher winter NPP/lower summer NPP in the south, timing differences in NPP seasonal phenology, and different NPP depth distribution patterns in the summer months. With a 6-month record of concurrent oxygen and bio-optical measurements from two Argo floats, the authors also demonstrated the ability of Argo profiling floats to obtain estimates of the net community production (NCP) to NPP ratio (f-ratio), ranging from 0.3 in July to -1.0 in December 2016.
This work highlights the utility of float bio-optical profiles in comparison to traditional measurements and indicates that environmental conditions (e.g. light availability, nutrient supply) are major factors controlling the seasonality and spatial (horizontal and vertical) distributions of NPP in the western North Atlantic Ocean.
Authors:
Bo Yang (University of Virginia, UM CIMAS/NOAA AOML)
James Fox (Oregon State University)
Michael J. Behrenfeld (Oregon State University)
Emmanuel S. Boss (University of Maine)
Nils Haëntjens (University of Maine)
Kimberly H. Halsey (Oregon State University)
Steven R. Emerson (University of Washington)
Scott C. Doney (University of Virginia)