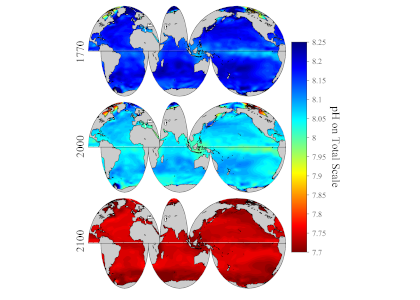
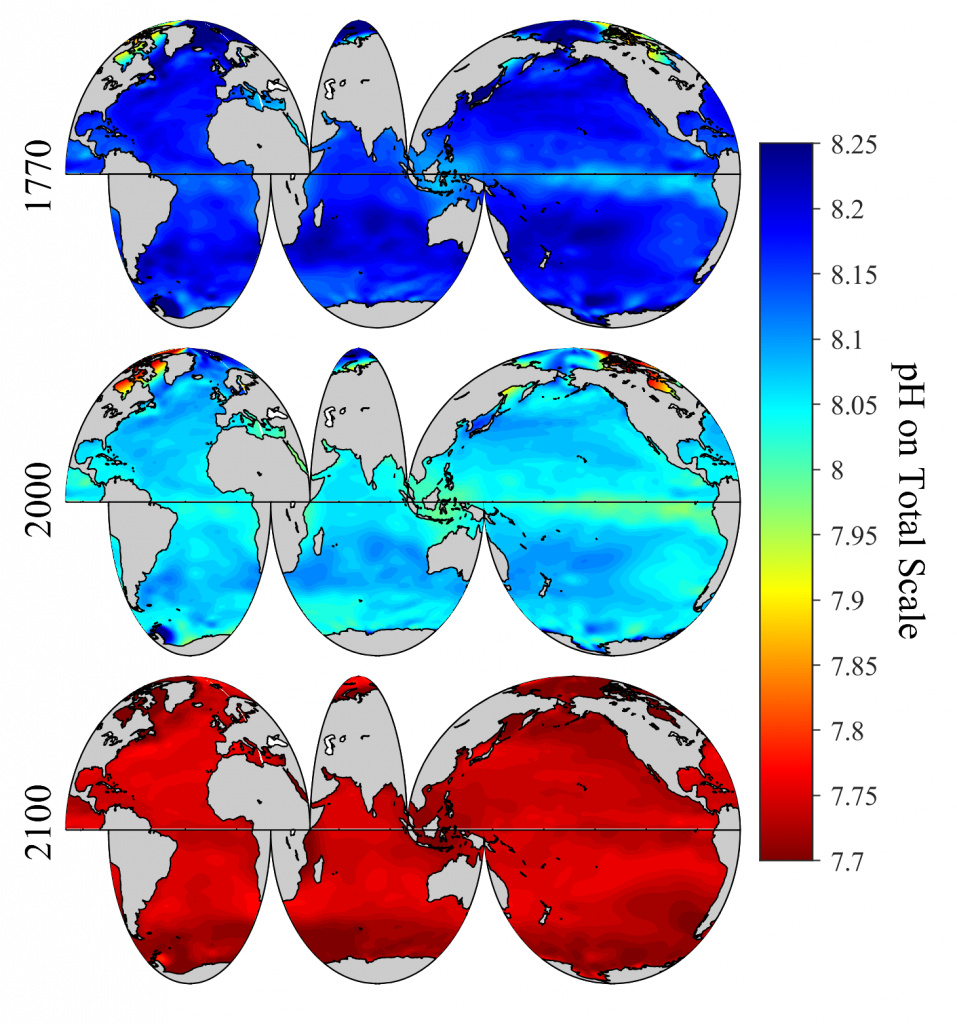
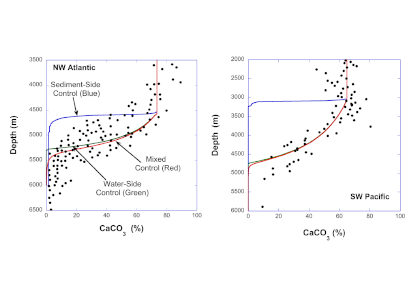
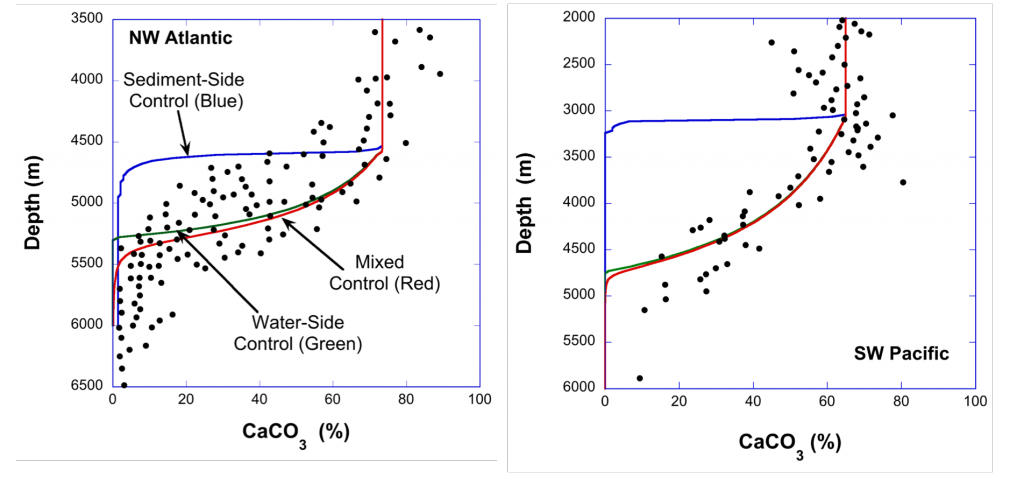
Vegetated coastal “blue carbon” ecosystems, including sea grasses, mangroves, and salt marshes, provide valuable ecosystem services such as carbon sequestration, storm protection, critical habitat, etc.. Many of these services are supported by the ability of blue carbon ecosystems to accumulate soil organic carbon over thousands of years. Rapidly changing climate and environmental conditions will impact decomposition and thus the global reservoir of organic carbon in coastal soils. A recent Perspective article published in Nature Geoscience focused on the biogeochemical factors affecting decomposition in coastal soils, such as mineral protection, redox zonation, water content and movement, and plant-microbe interactions. The authors explored the spatial and temporal scales of these decomposition mechanisms and developed a conceptual framework to characterize how they may respond to environmental disturbances such as land-use change, nutrient loading, warming, and sea-level rise.


Figure caption: Temperate salt marshes (MA, USA). Healthy salt marshes have lush stands of grasses (top). Storms can expose peat deposits that have been buried for thousands of years (bottom). The fate of this soil carbon is unknown, but some fraction will be respired by microbes and returned to the atmosphere as CO2.
Improved estimates of soil organic carbon in blue carbon systems will require better characterization of these processes from sustained data sets. Furthermore, incorporation of these decomposition mechanisms into ecosystem evolution models will improve our capacity to quantify and predict changes in these soil carbon reservoirs, which could facilitate their inclusion in global budgets and management tools.
Temperate salt marshes (MA, USA). Healthy salt marshes have lush stands of grasses (left/top). Storms can expose peat deposits that have been buried for thousands of years (right/bottom). The fate of this soil carbon is unknown, but some fraction will be respired by microbes and returned to the atmosphere as CO2.
Authors:
Amanda C Spivak (University of Georgia)
Jon Sanderman (Woods Hole Research Center)
Jennifer Bowen (Northeastern University)
Elizabeth A. Canuel (Virginia Institute of Marine Science)
Charles S Hopkinson (University of Georgia)