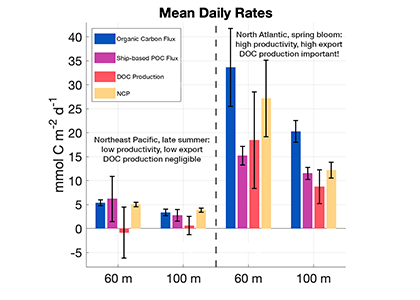
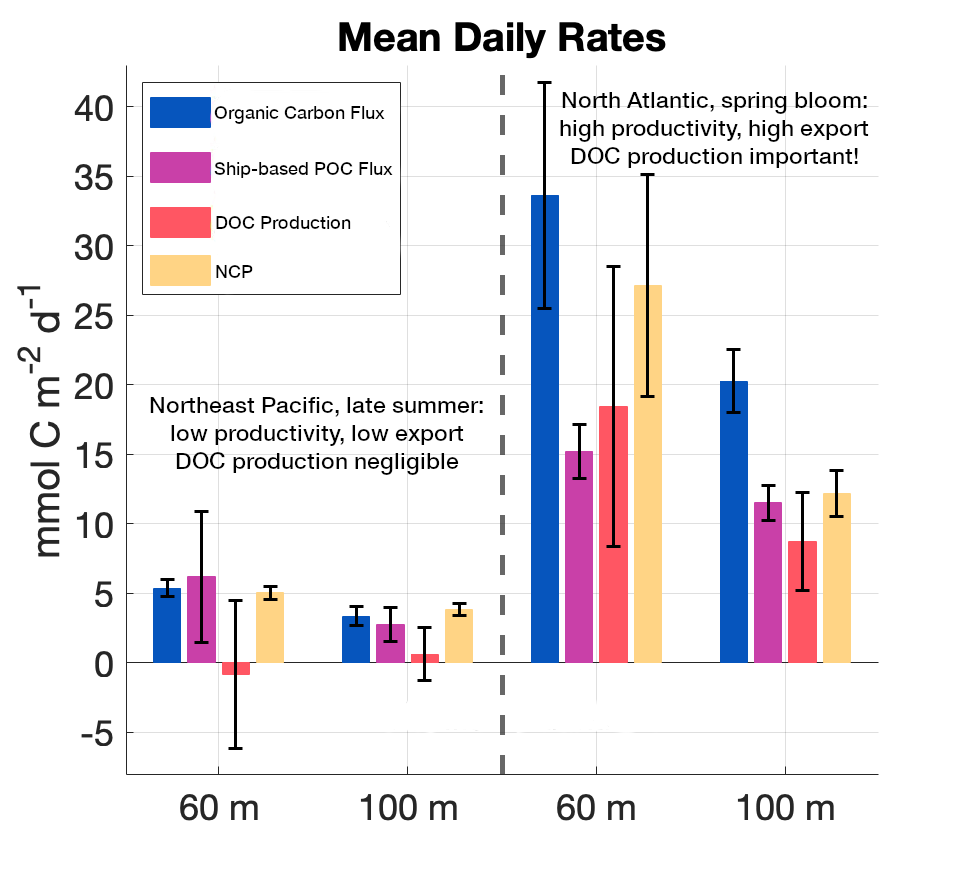
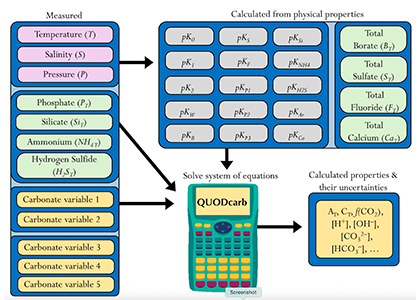
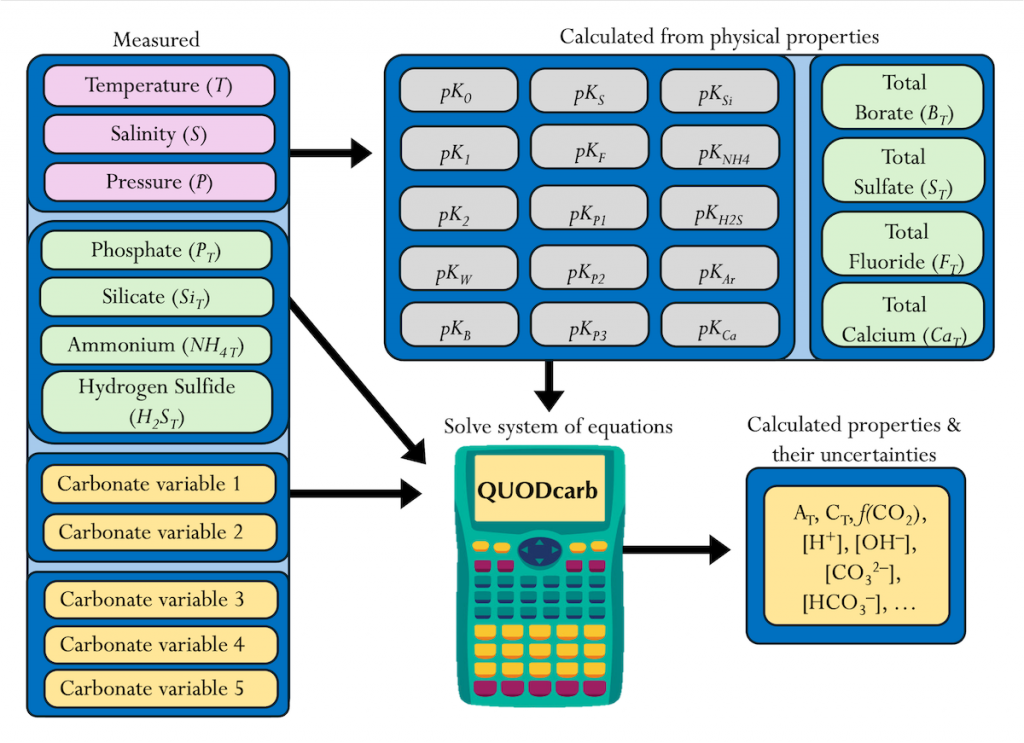
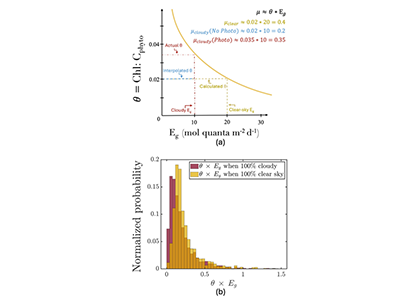
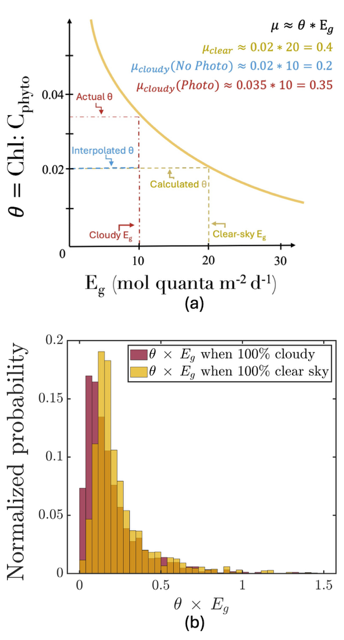
The SOLAS Summer School is a regular, international event with the goal to provide the multidisciplinary air-sea interaction background to the next generation of Earth System scientists. We are proud to celebrate the 10th SOLAS Summer School, with an expanded immersive three-week programme designed for early career researchers. Participants will explore the complex interactions between the ocean, atmosphere, and climate through a dynamic blend of lectures, hands-on practicals, collaborative student-led projects, all aligned with the SOLAS 3.0 framework “SOLAS Science Towards Solutions.”
SOLAS Summer School 2026 will take place from 9-27 March 2026 at Centro Nacional de Pesquisa e Conservação da Biodiversidade Marinha do Nordeste (CEPENE) in Tamandaré, Pernambuco (PE), Brazil.
To apply for Summer School 2026, please submit your application here by 31 August 2025. For students without access to Google Forms please download the form and email your application to solasschool@xmu.edu.cn.
Please send an email to solasschool@xmu.edu.cn, if you have any questions or need further assistance regarding the Summer School.