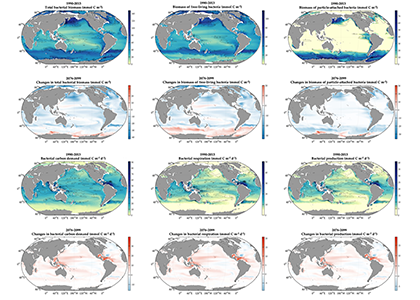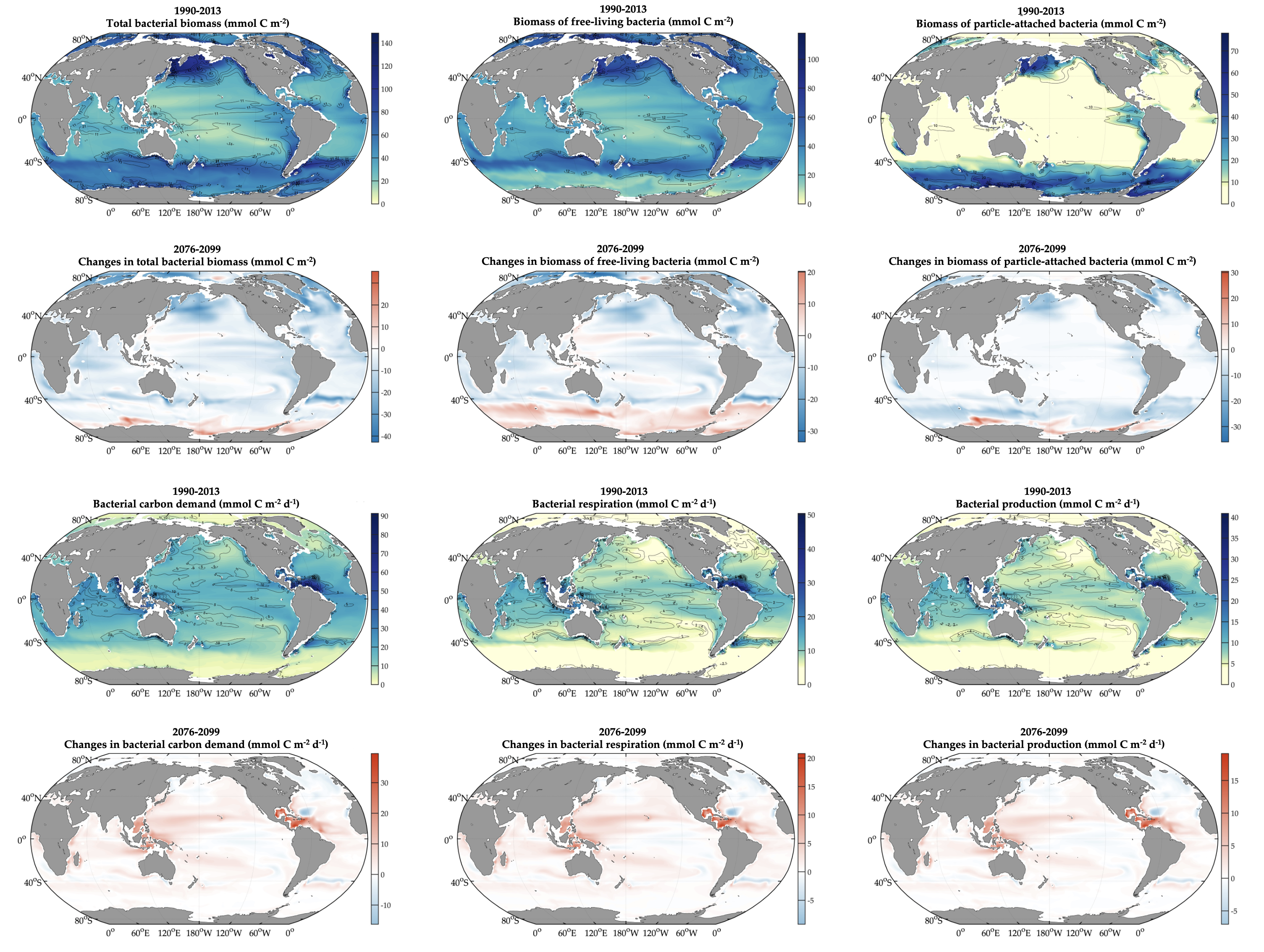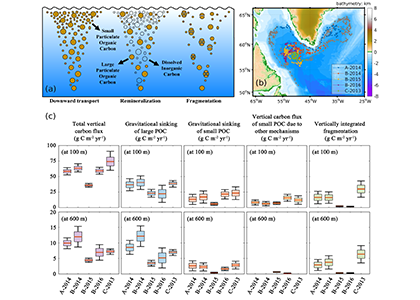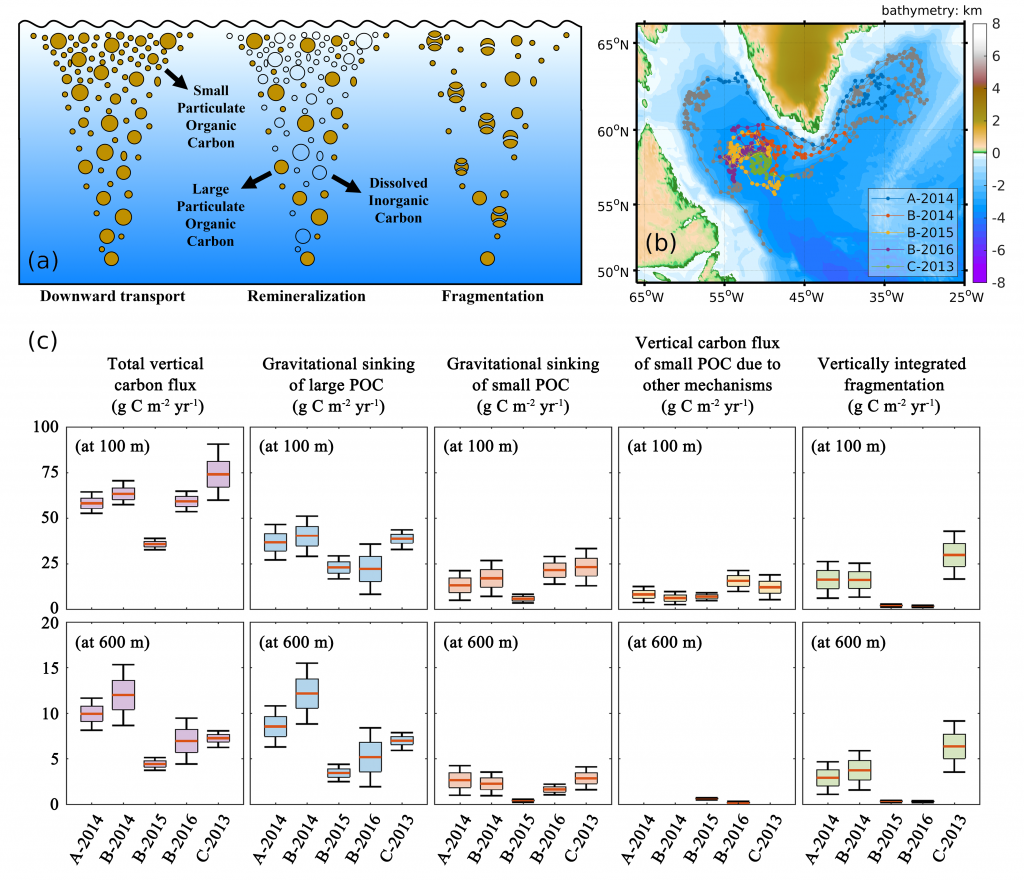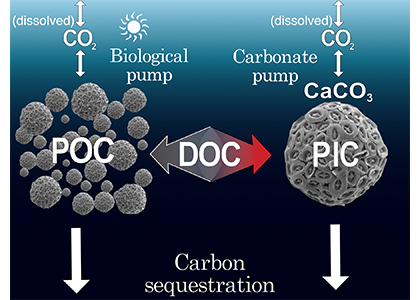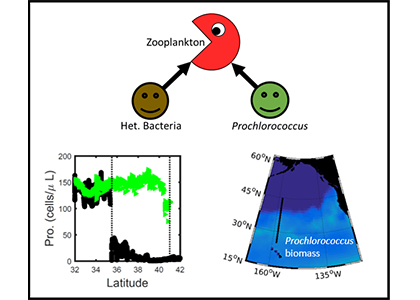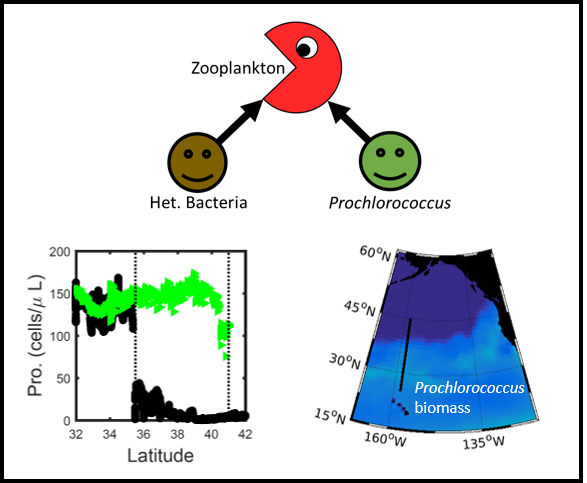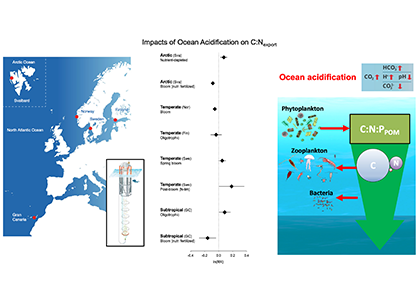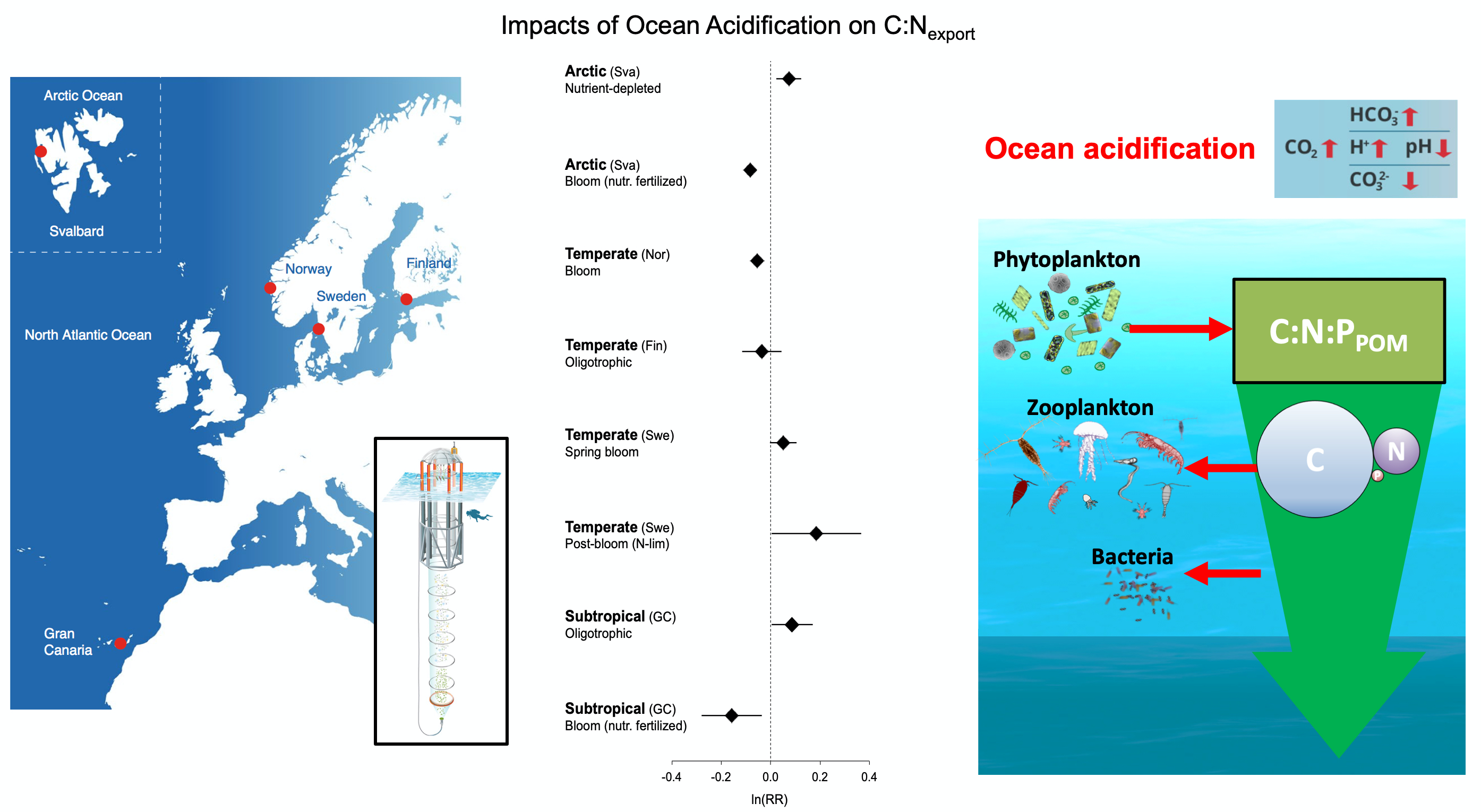
Deep sea microbial communities are experiencing increasing hydrostatic pressure with depth. It is known that some deep sea microbes require high hydrostatic pressure for growth, but most measurements of deep-sea microbial activity have been performed under atmospheric pressure conditions.
In a recent paper published in Nature Geoscience, the authors used a new device coined ‘In Situ Microbial Incubator’ (ISMI) to determine prokaryotic heterotrophic activity under in situ conditions. They compared microbial activity in situ with activity under atmospheric pressure at 27 stations from 175 to 4000 m depths in the Atlantic, Pacific, and the Southern Ocean. The bulk of heterotrophic activity under in situ pressure is always lower than under atmospheric pressure conditions and is increasingly inhibited with increasing hydrostatic pressure. Single-cell analysis revealed that deep sea prokaryotic communities consist of a small fraction of pressure-loving (piezophilic) microbes while the vast majority is pressure-insensitive (piezotolerant). Surprisingly, the piezosensitive fraction (~10% of the total community) responds with a more than 100-fold increase of activity upon depressurization. In the microbe proteomes, the authors uncovered taxonomically characteristic survival strategies in meso- and bathypelagic waters. These findings indicate that the overall heterotrophic microbial activity in the deep sea is substantially lower than previously assumed, which implies major impacts on the carbon budget of the ocean’s interior.
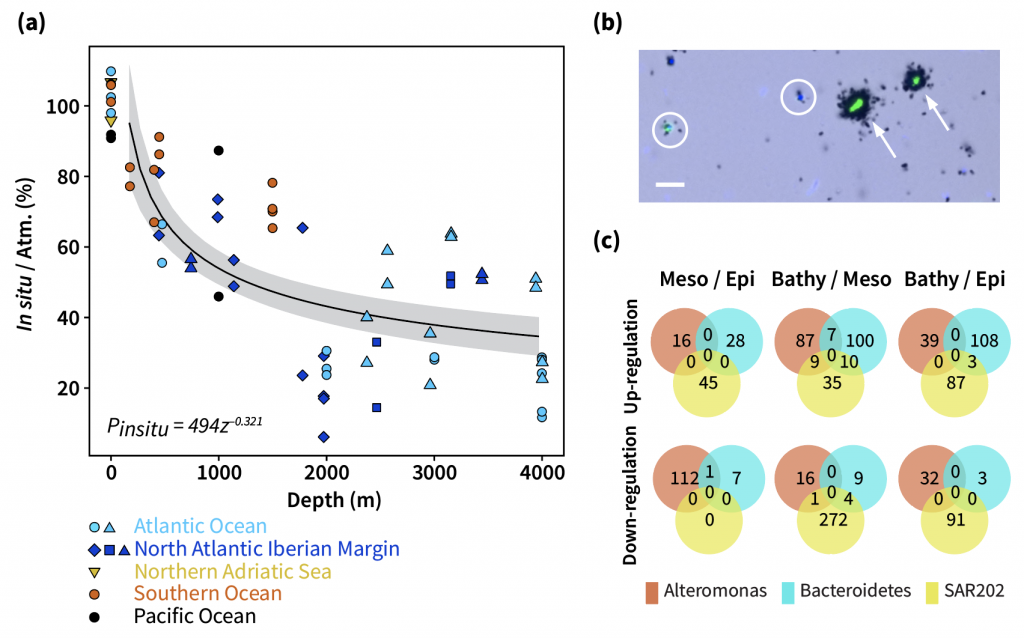
Figure caption: Deep sea microbial activity under varying pressure. (a) In situ bulk leucine incorporation rates normalized to rates obtained at atmospheric pressure conditions. (b) A microscopic view of a 2000 m sample collected in the Atlantic and incubated under atmospheric pressure conditions. The black halos around the cells are silver grains corresponding to their activities. The highly active cells (indicated by arrows) were rarely found in in situ pressure incubations. (c) Depth-related changes in the metaproteome of three abundant deep sea bacterial taxa (Alteromonas, Bacteroidetes, and SAR202). The number indicates shared and unique up- and down-regulated proteins in different depth zones.
Authors
Chie Amano (University of Vienna, Austria)
Zihao Zhao (University of Vienna, Austria)
Eva Sintes (University of Vienna, IEO-CSIC, Spain)
Thomas Reinthaler (University of Vienna, Austria)
Julia Stefanschitz (University of Vienna, Austria)
Murat Kisadur (University of Vienna, Austria)
Motoo Utsumi (University of Tsukuba, Japan)
Gerhard J. Herndl (University of Vienna, Netherlands Institute for Sea Research)
Twitter @microbialoceanW