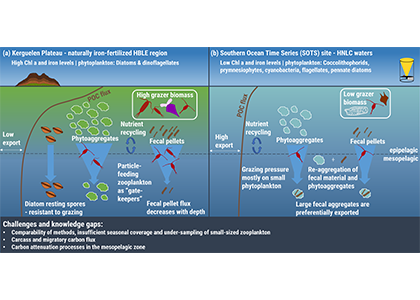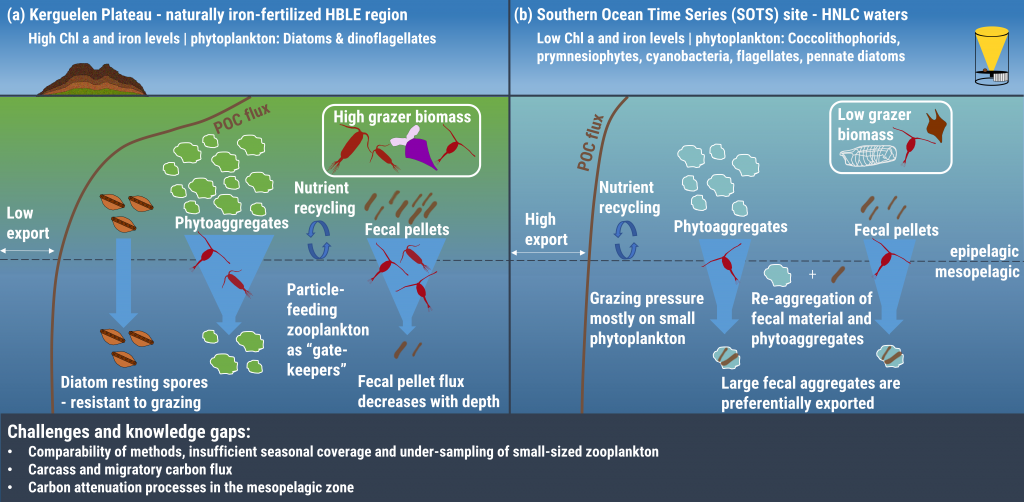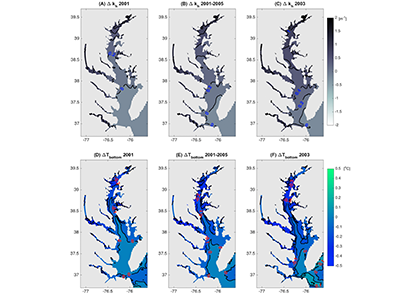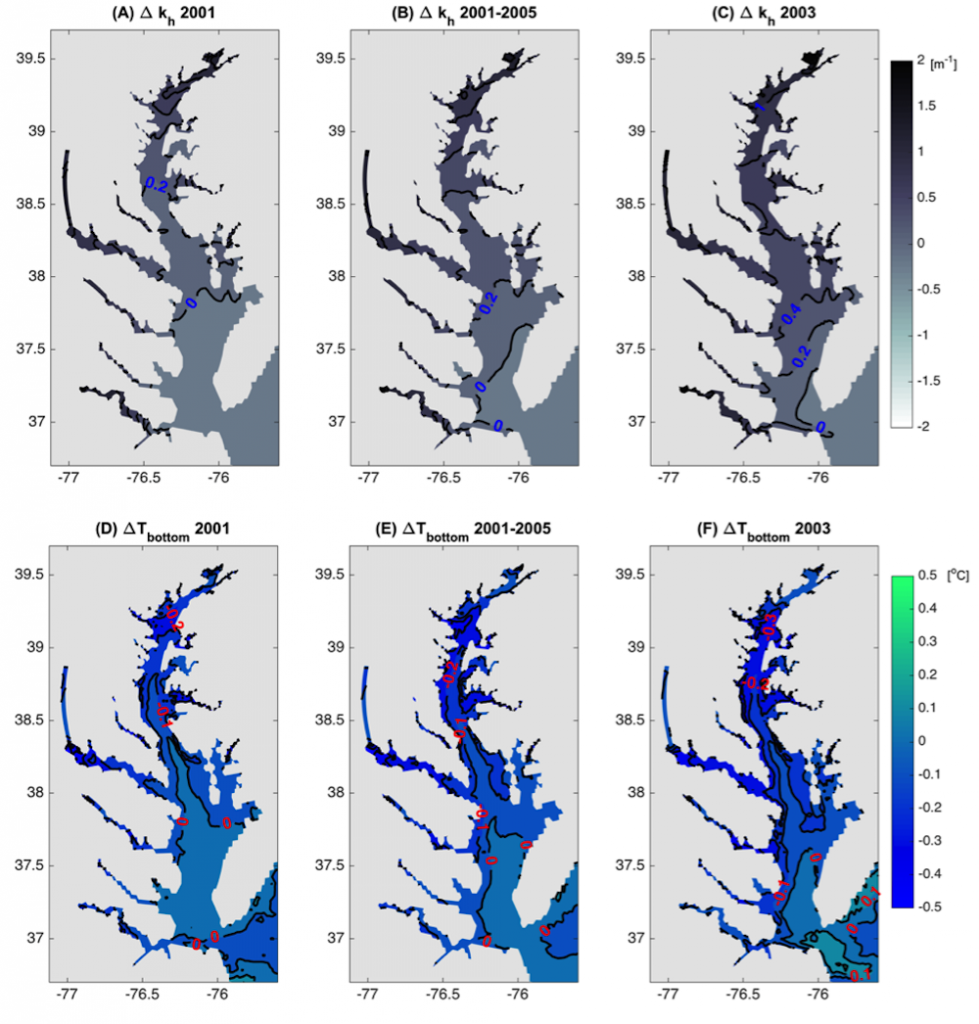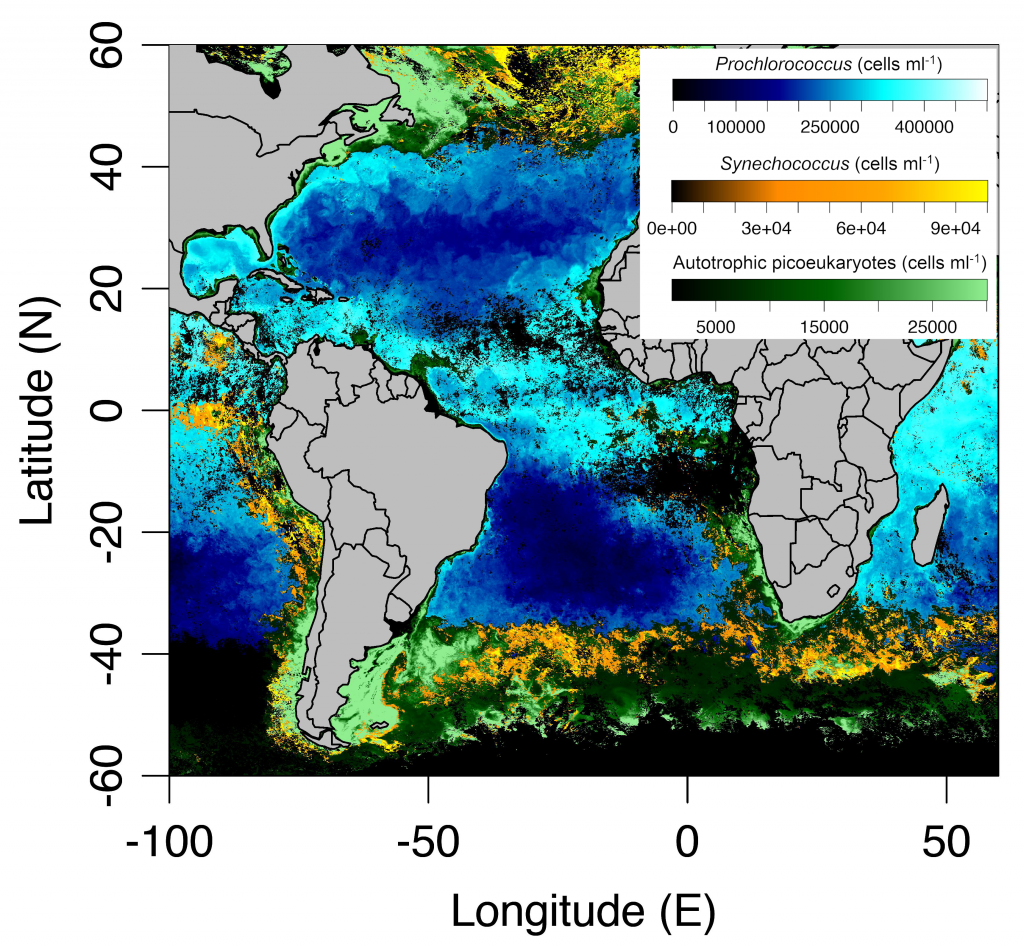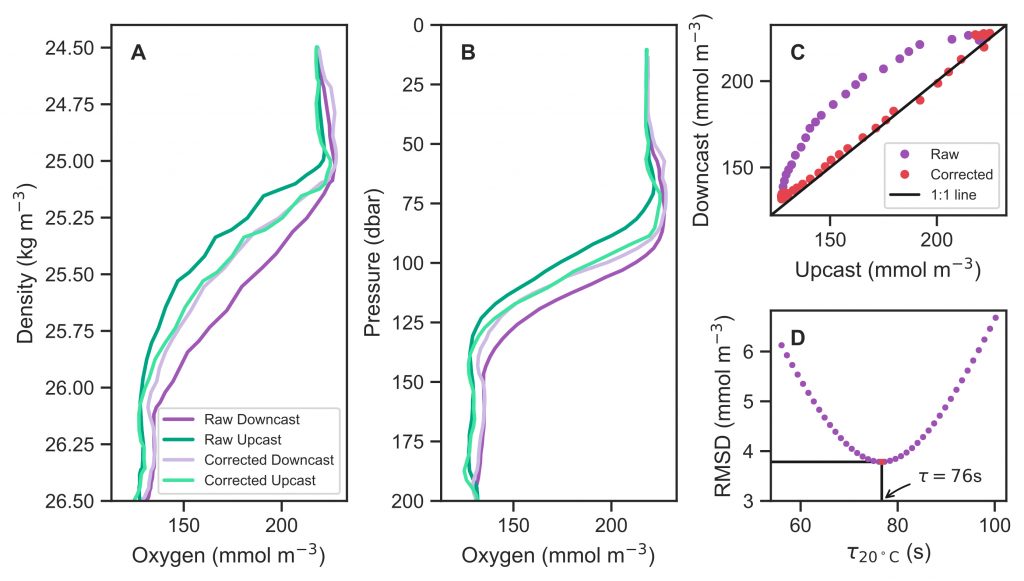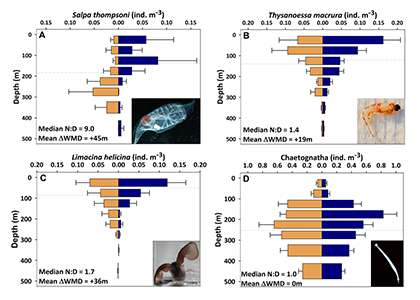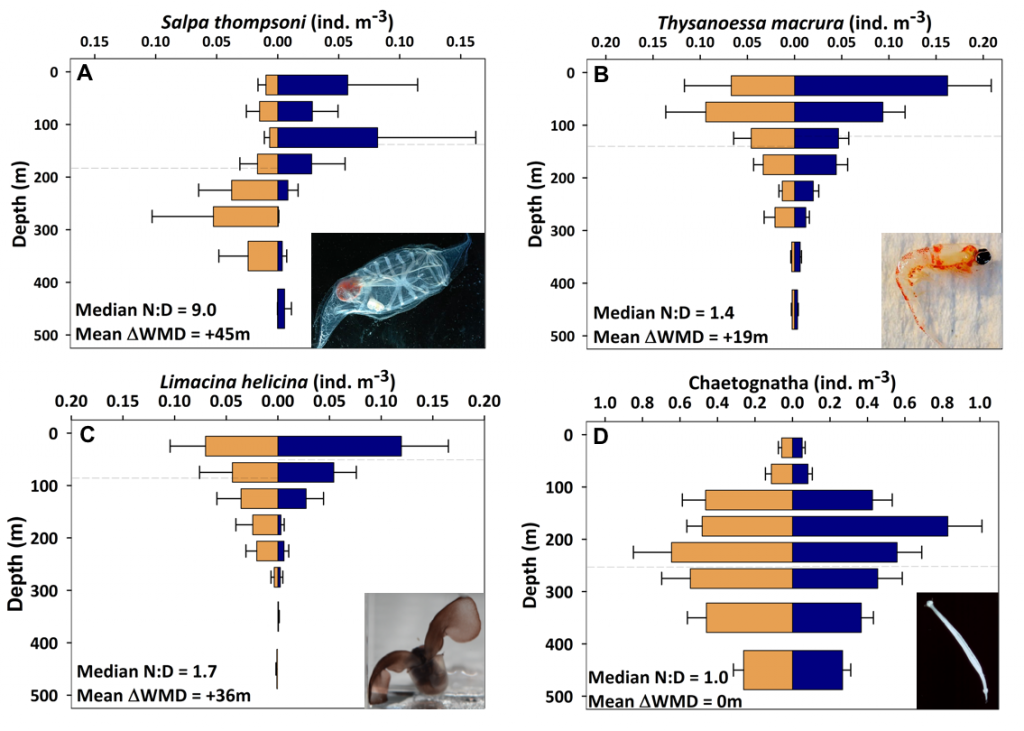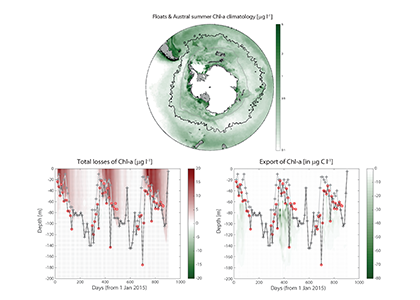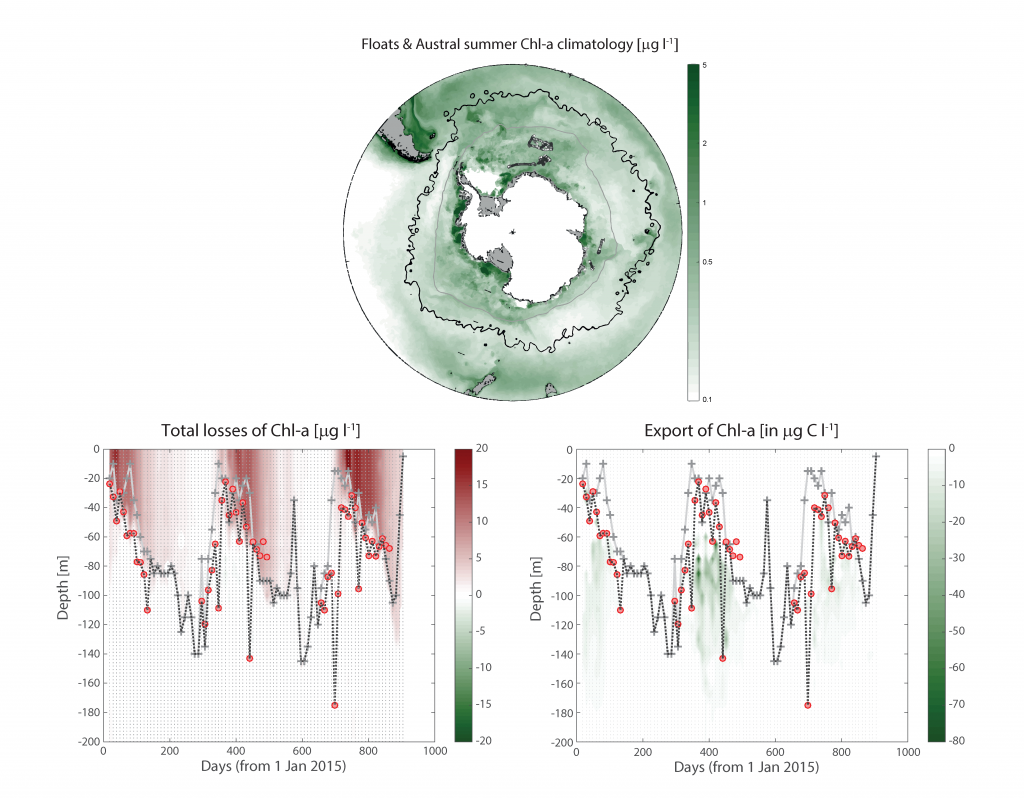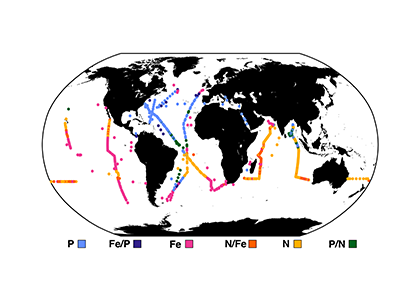
In the surface ocean, phytoplankton growth is often limited by a scarcity of key nutrients such as nitrogen, phosphorus, and iron. While this is important, there are methodological and conceptual difficulties in characterizing these nutrient limitations.
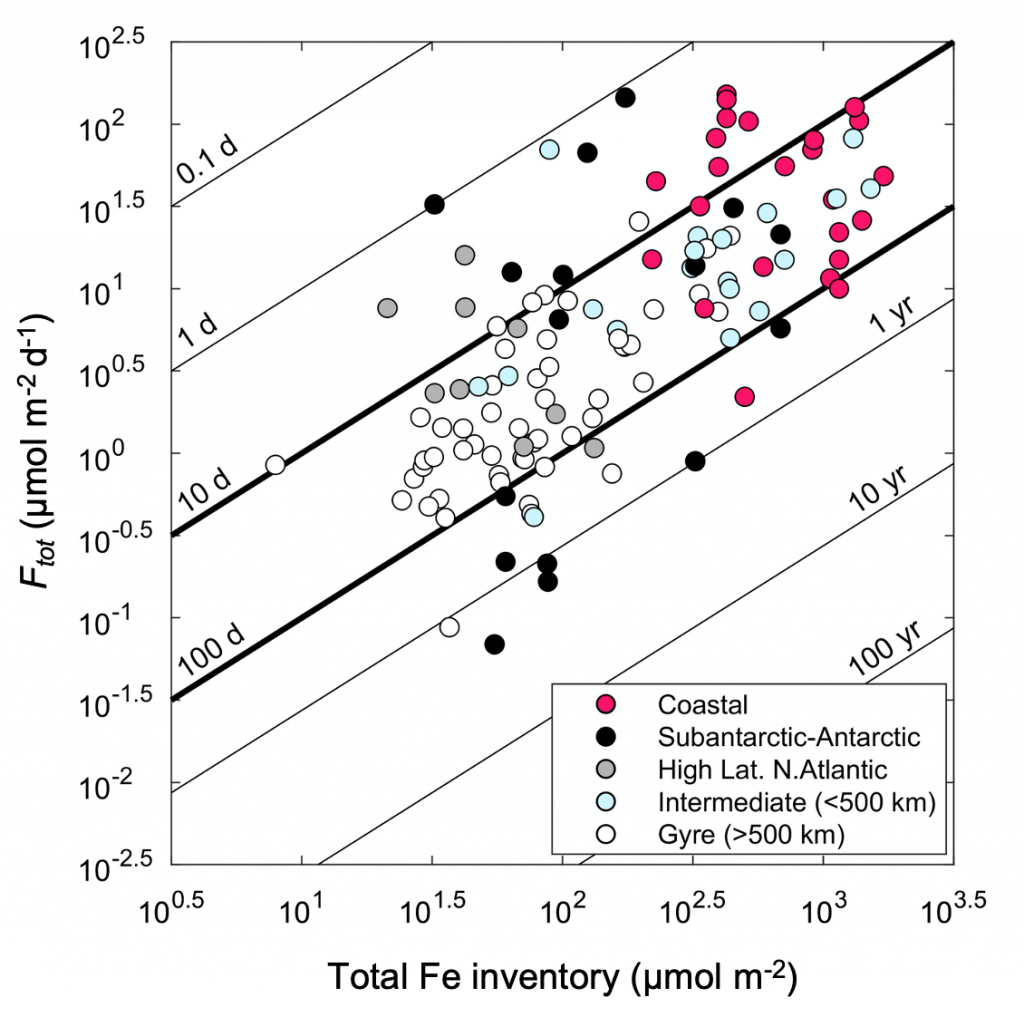
A recent paper published in Science Magazine leveraged a global metagenomic dataset from Bio-GO-SHIP to address these challenges. The authors characterized the abundance of genes that confer adaptations to nutrient limitation within the picocyanobacteria Prochlorococcus. Using the relative abundance of these genes as an indicator of nutrient limitation allowed the authors to capture expected regions of nutrient limitation, and novel regions that had not previously been studied. This gene-derived indicator of nutrient limitation matched previous methods of assessing nutrient limitation, such as bottle incubation experiments.
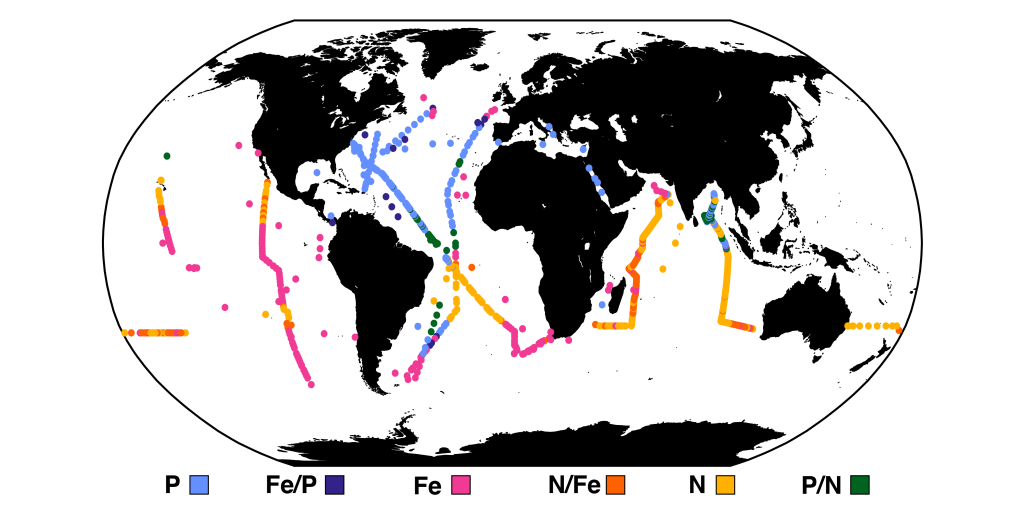
These findings have important implications for the global ocean. Characterizing the impact of nutrient limitation on primary production is especially critical in light of future stratification driven by climate change. In addition, this novel methodological approach allows scientists to use microbial communities as an eco-genomic biosensor of adaptation to changing nutrient regimes. For instance, future studies of coastal microbes or other ecosystems may help communities and environmental managers better understand how local microbial populations are adapting to climate change.
Watch an illustrated video overview of this research
Authors:
Lucas J. Ustick, Alyse A. Larkin, Catherine A. Garcia, Nathan S. Garcia, Melissa L. Brock, Jenna A. Lee, Nicola A. Wiseman, J. Keith Moore, Adam C. Martiny
(all University of California, Irvine)