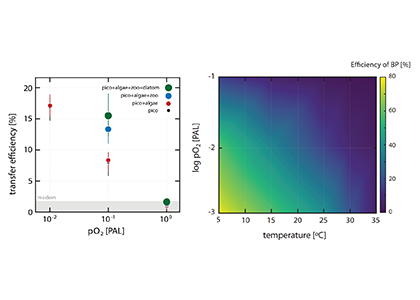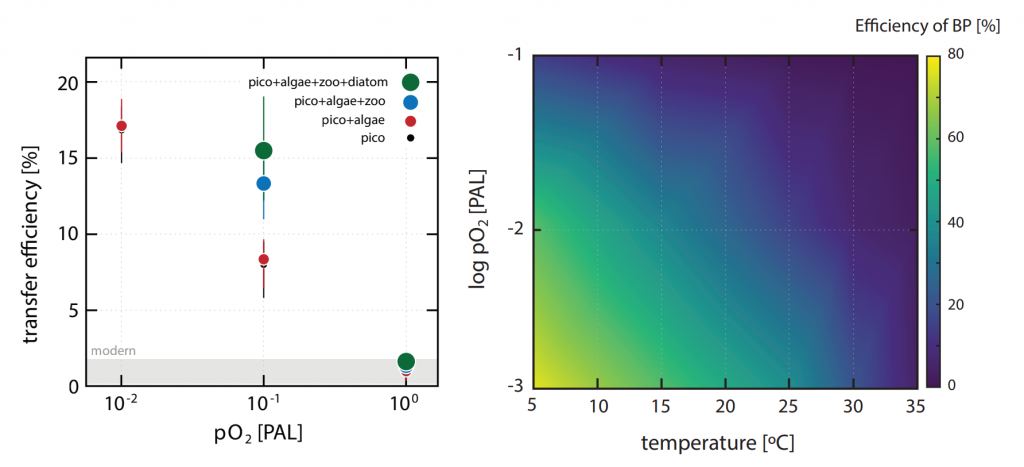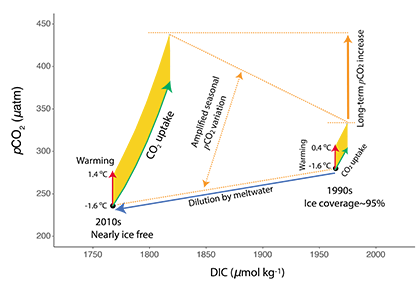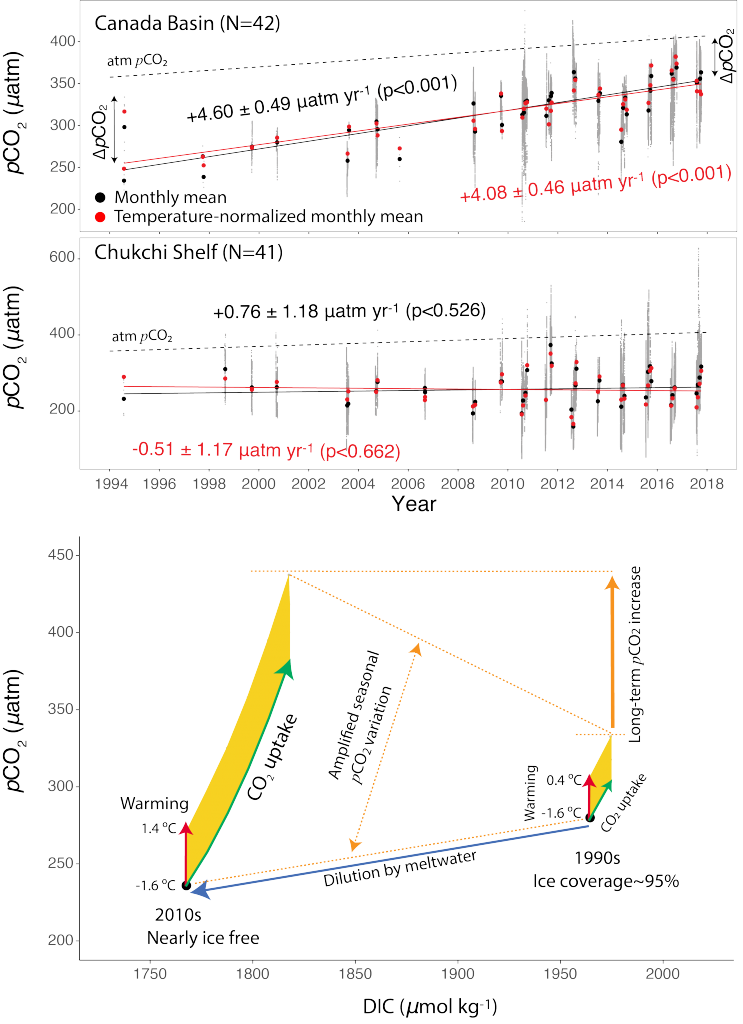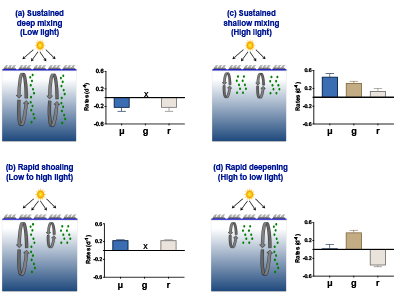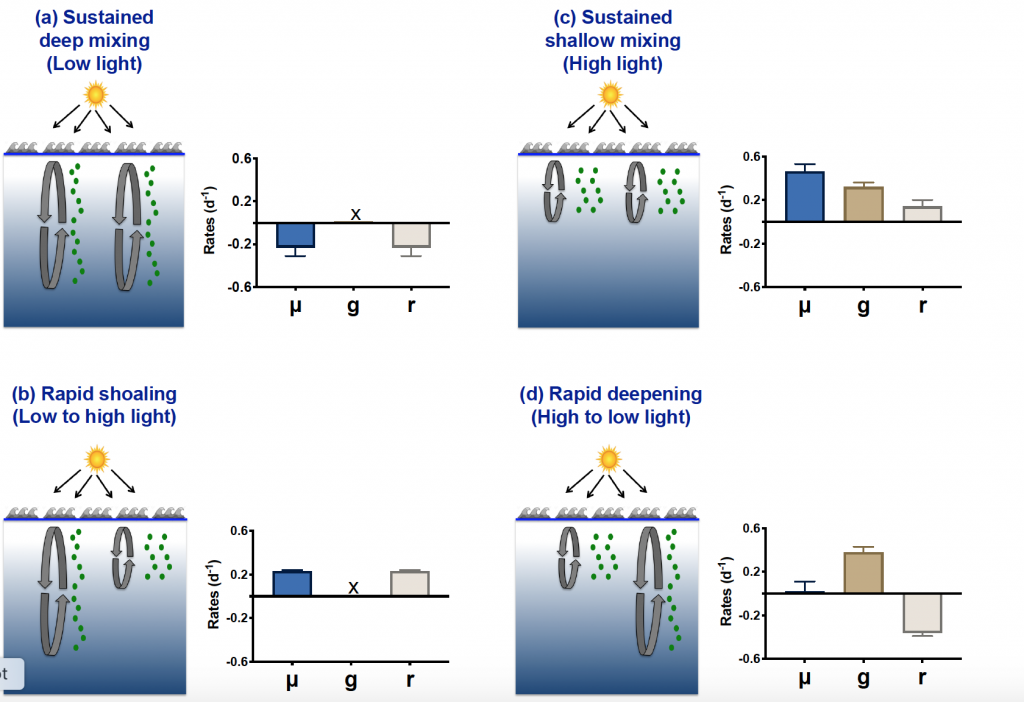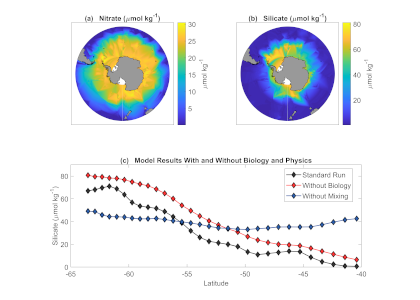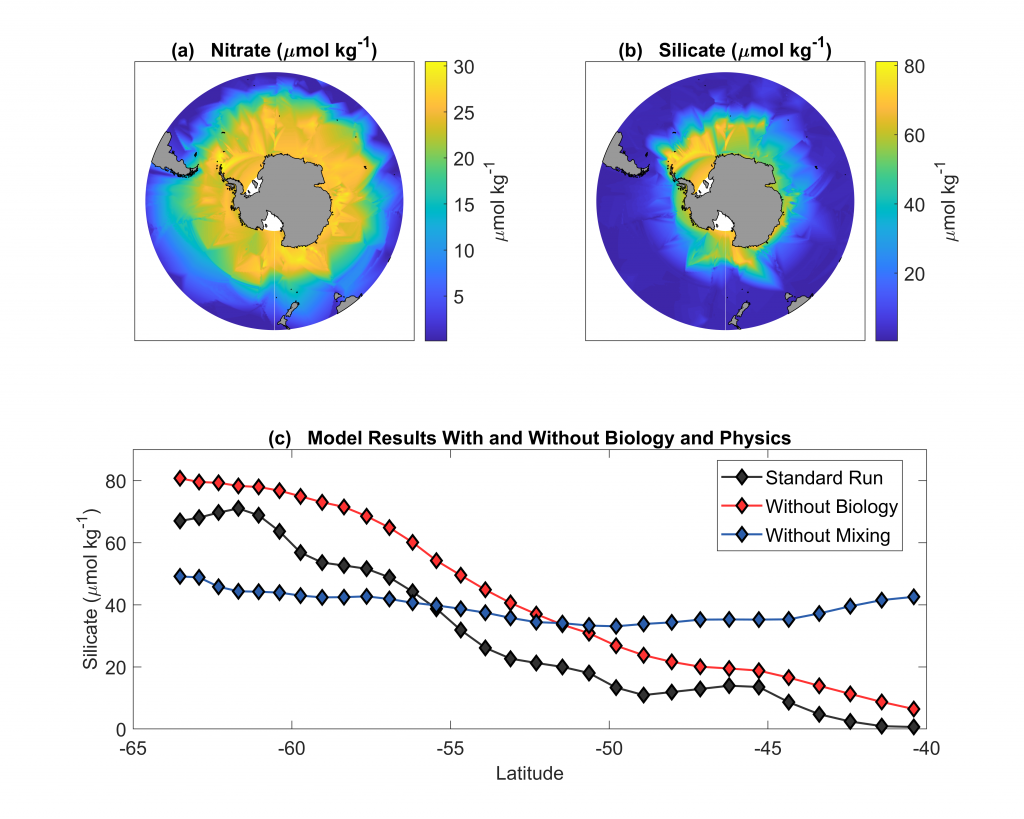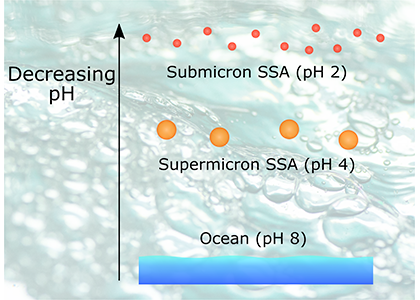
The pH of aerosols controls their impact on climate and human health. Sea spray aerosols are one of the largest sources of aerosols globally by mass, yet it has been challenging to measure the pH of fresh sea spray aerosols in the past. A recent study published in PNAS measured sea spray aerosols under controlled conditions, during a sampling intensive called SeaSCAPE, and optimized a pH paper-based technique to measure the aerosol acidity. The authors found that fresh sea spray aerosols can be rapidly acidified by 4 to 6 orders of magnitude relative to the ocean. This acidification is caused by interaction with surrounding acidic gases, changes in relative humidity, and enhanced dissociation of organic acids within the aerosols. This is a critical finding since the pH of aerosols controls key atmospheric chemical reactions including sulfur dioxide oxidation to form particulate sulfate. The results are also important in light of the fact that enzyme activity has been observed in sea spray aerosols, and enzyme activity is pH dependent.
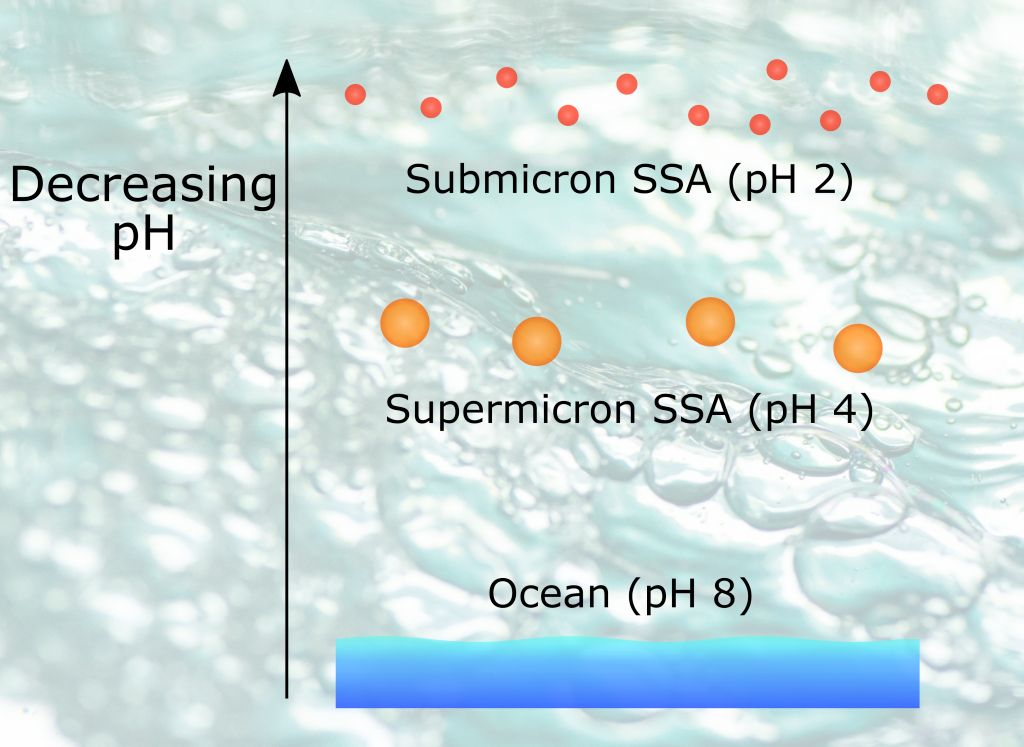
Figure 1. Acidity of nascent sea spray aerosols (SSA) compared to bulk ocean water measured during the 2019 SeaSCAPE sampling intensive. Background artwork by Nigella Hillgarth.
Authors
Kyle Angle (University of California, San Diego)
Daniel Crocker (University of California, San Diego)
Rebecca Simpson (University of California, San Diego)
Kathryn Mayer (University of California, San Diego)
Lauren Garofalo (Colorado State University, Fort Collins)
Alexia Moore (University of California, San Diego)
Stephanie Mora Garcia (University of California, San Diego)
Victor Or (University of California, San Diego)
Sudarshan Srinivasan (University of California, San Diego)
Mahum Farhan (University of California, San Diego)
Jonathan Sauer (University of California, San Diego)
Christopher Lee (University of California, San Diego)
Matson Pothier (Colorado State University, Fort Collins)
Delphine Farmer (Colorado State University, Fort Collins)
Todd Martz (University of California, San Diego)
Timothy Bertram (University of Wisconsin, Madison)
Christopher Cappa (University of California, Davis)
Kimberly Prather (University of California, San Diego)
Vicki Grassian (University of California, San Diego)
Joint post with Surface Ocean – Lower Atmosphere Study (SOLAS)