
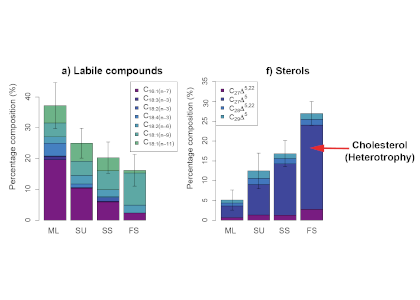
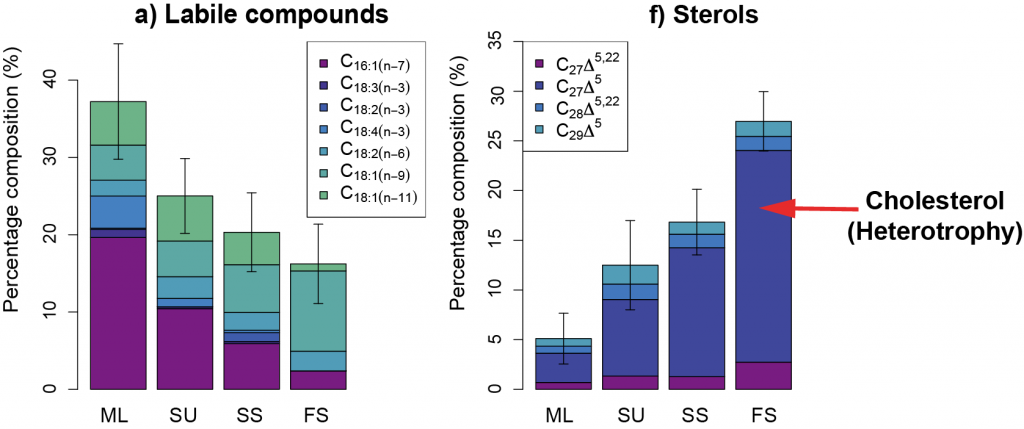
The Southern Ocean is iron (Fe)-limited, and increased fluxes of dust-borne Fe to the Southern Ocean during the Last Glacial Maximum (LGM) have been associated with phytoplankton growth and CO2 drawdown. Dust contains different mixes of Fe-bearing minerals, depending on the source region. Fe(II) silicate minerals from physical weathering are more bioavailable than Fe(III) oxyhydroxide minerals from chemical weathering. The Fe(II) silicates are dominant in dust sources that have been weathered from bedrock by glaciers in Patagonia, but the impact of glacial activity on dust-borne Fe speciation (Fe oxidation state and mineral composition) and bioavailability over the last glacial cycle has not previously been quantified.
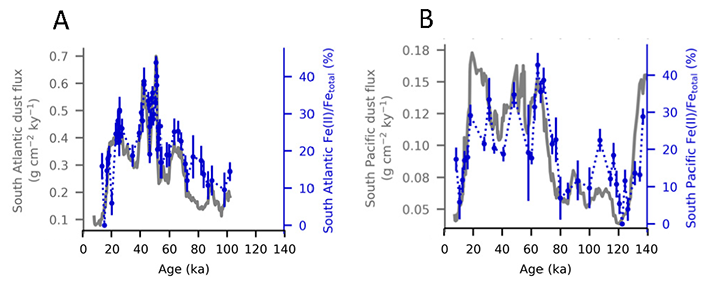
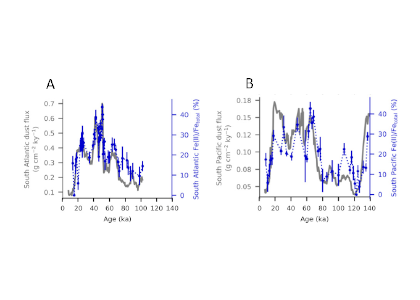
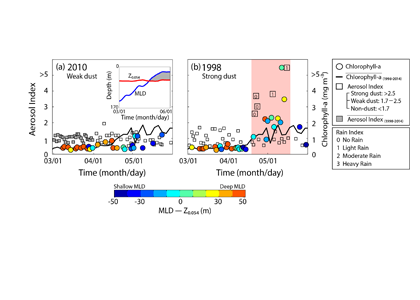
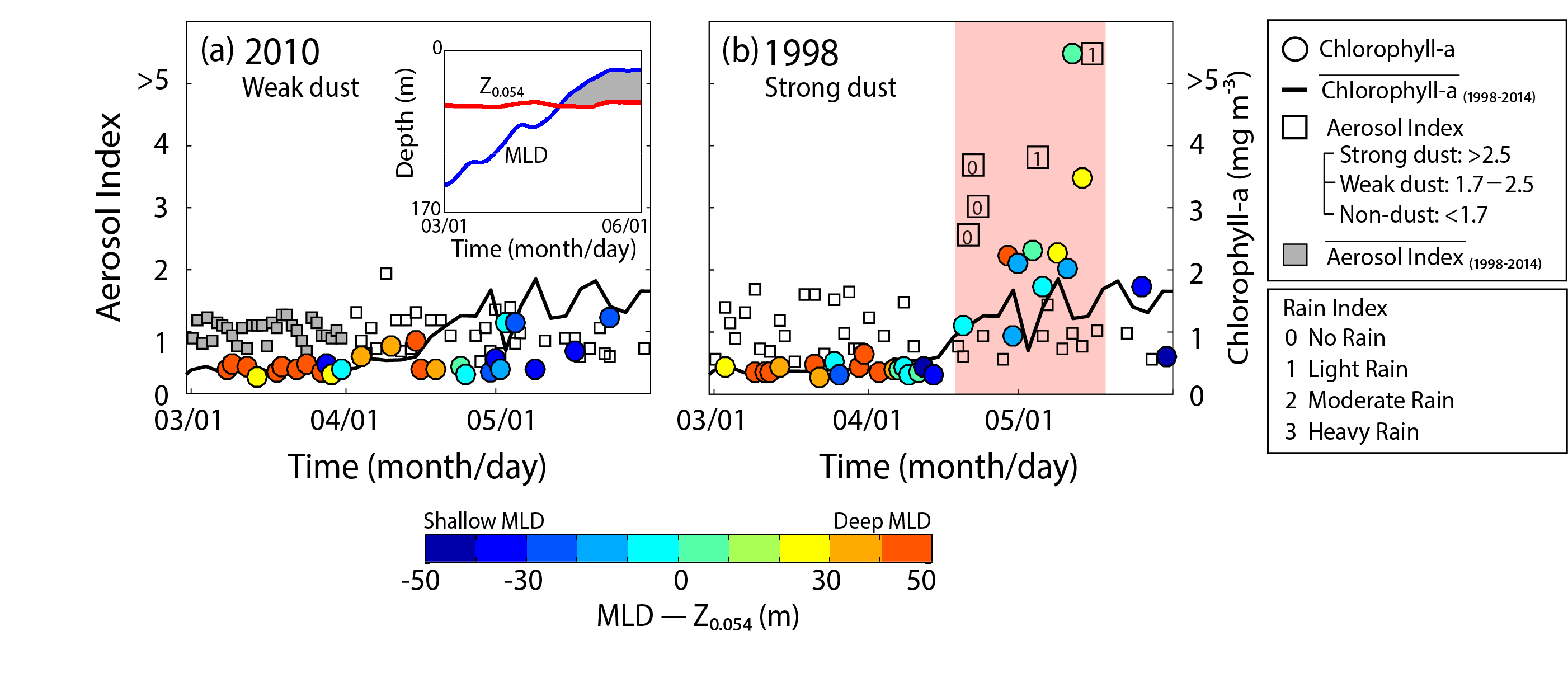
Figure 1. The fraction of Fe(II) in dust (Fe(II)/Fetotal, dominated by Fe(II) silicates, shown as blue dots connected with dotted lines on blue axes) in marine sediment cores from (A) the South Atlantic and (B) the South Pacific plotted with the total dust flux (grey lines on grey axes).
A recent study in PNAS reconstructs the speciation of dust-borne Fe over the last glacial cycle in South Atlantic and South Pacific marine sediment cores using Fe K-edge X-ray absorption spectroscopy. The authors observed that the highly bioavailable Fe(II) silicate content of dust-borne Fe is higher in both regions during cold glacial periods, suggesting that a given flux of Fe is more bioavailable in glacial versus interglacial periods (Figure 1). Therefore, all Fe cannot be considered equal in biogeochemical models working on glacial-interglacial timescales. The bioavailability of a given flux of Fe at the LGM was likely a dominant driver of phytoplankton growth, with more bioavailable Fe driving increased phytoplankton activity and associated atmospheric CO2 drawdown and subsequent cooling. The observed association between glacial periods and increased Fe bioavailability in the Southern Ocean may indicate an important positive feedback mechanism between glacial activity and cold glacial temperatures through Fe speciation and the efficiency of the biological pump.
Paper link: https://doi.org/10.1073/pnas.1809755115
Authors:
Elizabeth M. Shoenfelt (Lamont-Doherty Earth Observatory, Columbia University)
Gisela Winckler (Lamont-Doherty Earth Observatory, Columbia University)
Frank Lamy (Alfred Wegener Institute, Helmholtz Centre for Polar and Marine Research)
Robert F. Anderson (Lamont-Doherty Earth Observatory, Columbia University)
Benjamin C. Bostick (Lamont-Doherty Earth Observatory, Columbia University)



{kind=link}