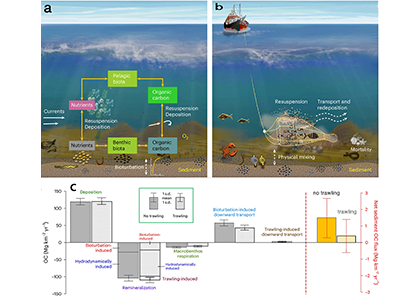
Bottom trawling, a fishing method that uses heavy nets to catch animals that live on and in the seafloor, could release a large amount of organic carbon from seafloor into the water, that metabolizes to CO2 then outgasses to the atmosphere. The magnitude of this indirect emission has been heavily debated, with estimates spanning from negligibly small to global climate relevant. Thus, a lack of reliable data and insufficient understanding of the process hinders management of bottom trawling for climate protection.
We set out to solve this problem in two steps. First, we analyzed a large field dataset containing more than 2000 sediment samples from one of the most intensely trawled regions globally, the North Sea. We identified a trawling-induced carbon reduction trend in the data, but only in samples taken in persistently intensively trawled areas with multi-year averaged swept area ratio larger than 1 yr-1. In less intensely trawled areas, there was no clear effect. In a second step, we applied numerical modelling to understand the processes behind the observed change (Fig. 1). Our model results suggest that bottom trawling annually releases one million tonnes of CO2 in the North Sea and 30 million tonnes globally. Along with sediment resuspension in the wake of the trawls, the main cause for altered sedimentary carbon storage is the depletion of macrofauna, whose locomotion and burrowing effectively buries freshly deposited carbon into deeper sediment layers. By contrast, macrofauna respiration is reduced owing to trawling-caused mortality, partly offsetting the organic carbon loss. Following a cessation of trawling, the simulated benthic biomass can recover in a few years, but the sediment carbon stock would take several decades to be restored to its natural state.
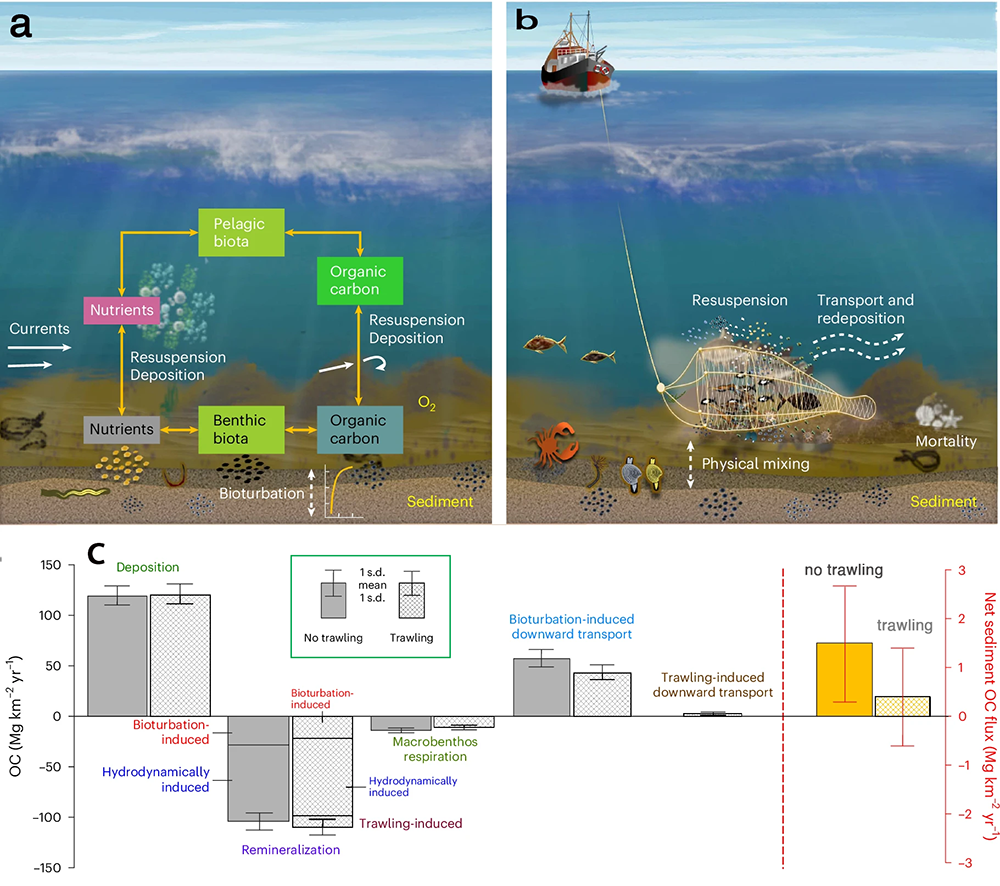
Figure 1. (a) Benthic–pelagic coupling in a natural system. (b) Processes involved in bottom trawling. (c) Model-estimated source and sink terms of organic carbon in surface sediments in the No-trawling (solid fill, n = 67 annual values for 1950–2016) and trawling (pattern fill, n = 67 ensemble-averaged values for 1950–2016) scenarios of the North Sea. © 2024, Zhang, W. et al., CC BY 4.0.
Marine conservation strategies traditionally favor hard bottoms, such as reefs, that are biologically diverse but accumulate limited amounts of organic carbon. Our results indicate that carbon in muddy sediments is more susceptible to trawling impacts than carbon in sand and point out a need to safeguard muddy habitats for climate protection. Our methods and results might be used in the context of marine spatial planning policies to gauge the potential benefits of limiting or ending bottom trawling within protected areas.
Zhang, W., Porz, L., Yilmaz, R. et al. Long-term carbon storage in shelf sea sediments reduced by intensive bottom trawling. Nat. Geosci. 17, 1268–1276 (2024). https://doi.org/10.1038/s41561-024-01581-4
Authors
Wenyan Zhang (Hereon)
Lucas Porz (Hereon)
Rümeysa Yilmaz (Hereon)
Klaus Wallmann (GEOMAR)
Timo Spiegel (GEOMAR)
Andreas Neumann (Hereon)
Moritz Holtappels (AWI)
Sabine Kasten (AWI)
Jannis Kuhlmann (BUND)
Nadja Ziebarth (BUND)
Bettina Taylor (BUND)
Ha Thi Minh Ho-Hagemann (Hereon)
Frank-Detlef Bockelmann (Hereon)
Ute Daewel (Hereon)
Lea Bernhardt (HWWI)
Corinna Schrum (Hereon)