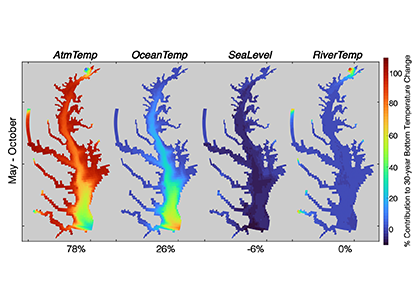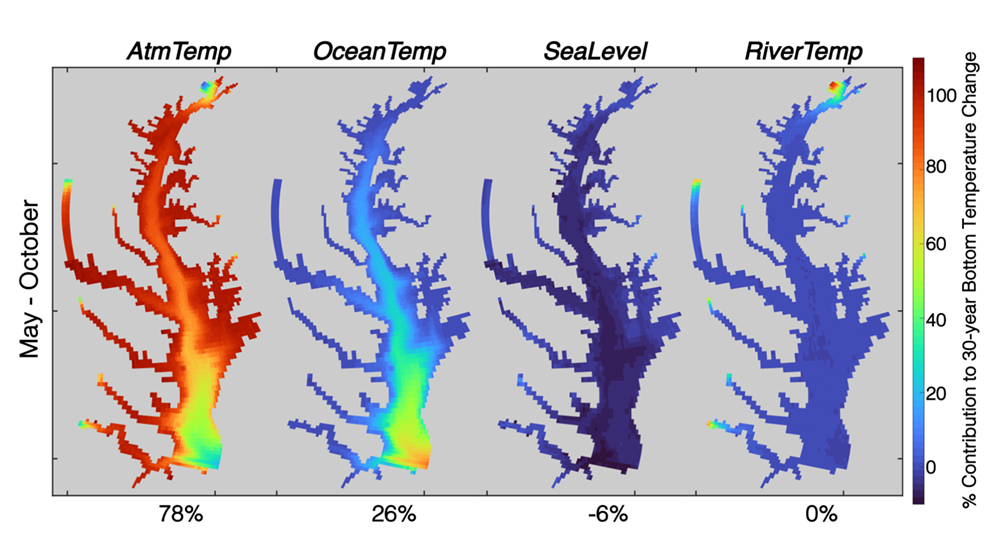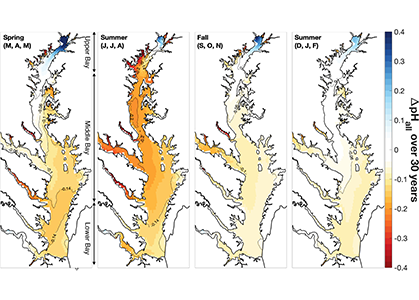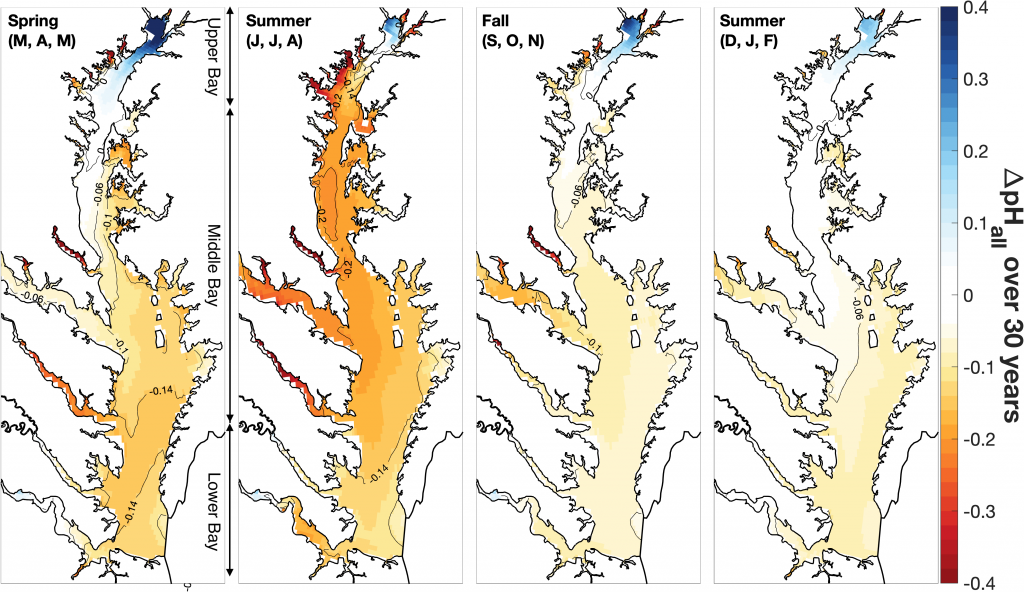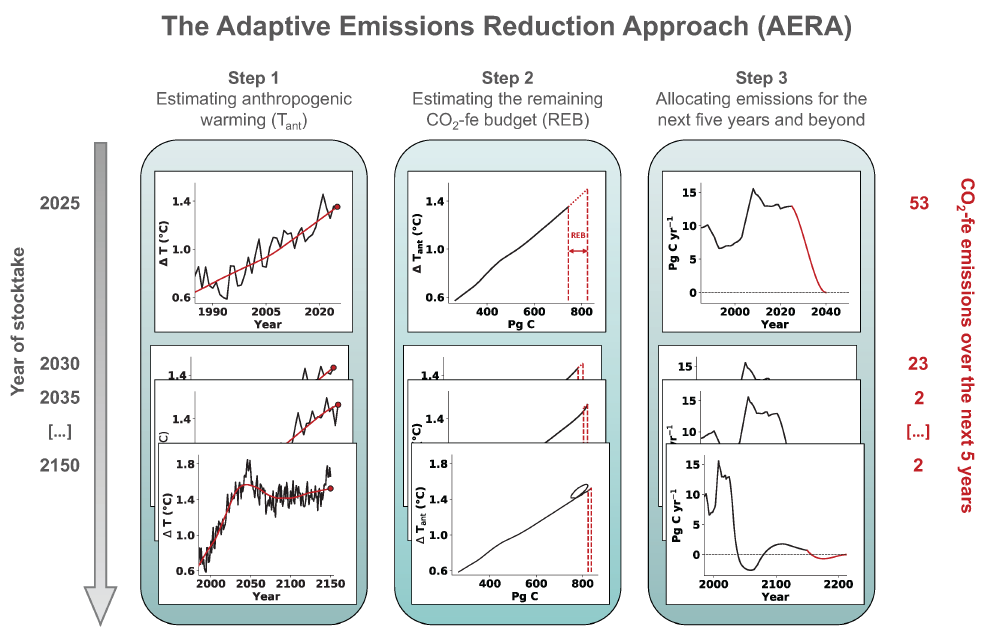
Around the world, countries have agreed in the Paris Agreement to limit global warming well below 2°C and to pursue efforts to reduce global warming to 1.5°C. However, large uncertainties remain about which emission pathways will allow us to reach this goal. A recent paper presents a new adaptive approach to create emission pathways and estimate the necessary emission reductions every five years, following the stocktake process of the Paris Agreement. This Adaptive Emissions Reduction Approach (AERA) is solely based on past warming rates, and emissions of CO2 and non-CO2 radiative agents, and explicitly does not rely on projections by Earth System Models. Updating the emission pathways every five years, circumvents uncertainties in the climate system and its transient response to cumulative emissions (TCRE). Testing with the Bern3D-LPX Earth System Model of Intermediate Complexity shows that the approach works robustly across a wide range of TCREs, avoids large overshoots, and only small changes to the emission pathways are necessary every five years. This approach will allow policymakers to estimate emission pathways and create a base for international negotiations. Furthermore, it allows simulations with Earth System Models that all converge to the same temperature target to compare the climate at stabilized warming levels.
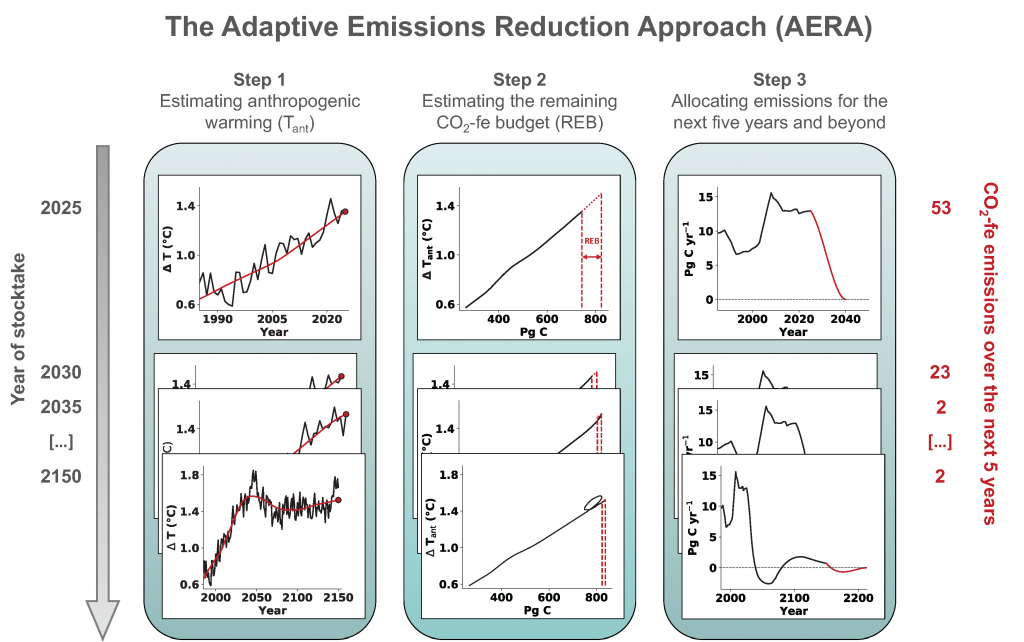
Figure caption: The three steps of the Adaptive Emission Reduction Approach: 1) Estimating the past anthropogenic warming, 2) estimating the remaining emission budget, and 3) redistributing it over the future years.
Authors
Jens Terhaar (University of Bern, now Woods Hole Oceanographic Institution)
Thomas L Frölicher (University of Bern)
Mathias T Aschwanden (University of Bern)
Pierre Friedlingstein (University of Exeter, Ecole Normale Superieure)
Fortunat Joos (University of Bern)
Twitter @JensTerhaar @froeltho @PFriedling @unibern @snsf_ch @4C_H2020 @ExeterUniMaths @Geosciences_ENS @IPSL_outreach