
We rely on global ocean models to predict how climate change might affect the evolution of ocean productivity, acidification, and deoxygenation (1). Such platforms are also used to test hypotheses regarding the controls on ocean biogeochemical cycling and to understand past change (both on historical and geologic timescales). Ocean biogeochemistry models began with relatively simple formulations of a carbon export flux that involved restoring to observed phosphate distributions, but have more recently evolved into complex multi-element representations of the ocean. In line with our understanding that the trace micronutrient iron (Fe) limits phytoplankton productivity over large areas of the world ocean (2), most global models that aim to project future change also explicitly represent the Fe cycle.
Datasets regarding the major limiting nutrients (nitrate, phosphate, and silicate) have been available as gridded ‘climatologies’ since the early 1990s (3). This has greatly facilitated the development and evaluation of modelled distributions over the past two decades. However, over this period there has been little comprehensive evaluation of how different models represent the ocean Fe cycle. Over recent years, there has been a marked increase in the availability of iron measurements in the ocean (4), largely driven by the international GEOTRACES effort to conduct full depth, basin-scale surveys. This led us to initiate the first-ever attempt to critically compare a range of global ocean iron models against the largest global datasets, as well as against the newly emerging ocean section data (5).
The Fe Model Intercomparison Project (FeMIP) sought to be as inclusive as possible in this first step and therefore did not seek to standardize the underlying ocean circulation or external inputs. Instead, we simply asked each of the thirteen models to provide their best representation of dissolved iron in three dimensions at monthly resolution. We then compared these models against each other, a global iron database of over 20,000 observations and against five unique basin-scale sections from the GEOTRACES intermediate data product 2014 (IDP2014) (6).
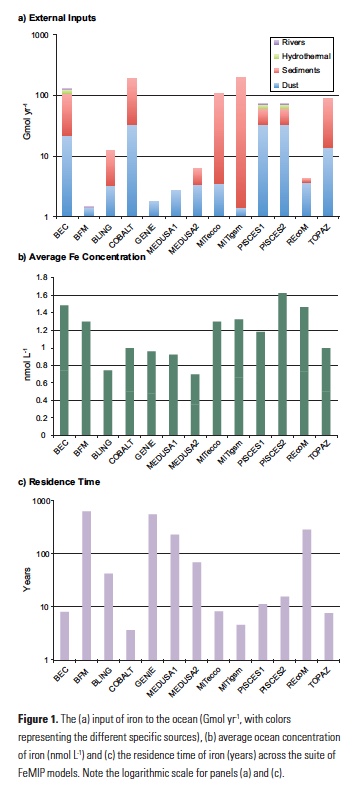
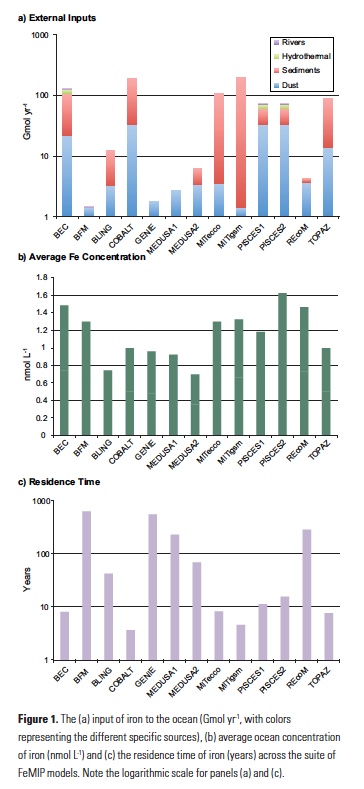
Firstly, it is apparent that even when the underlying iron cycles of the different models are evaluated, a substantial degree of inter-model discord exists. The total iron input varies from around 2 to 200 Gmol yr-1 across the thirteen models (Fig. 1a). Even for ‘well known’ sources like atmospheric deposition, the inter-model variability is around an order of magnitude. On the other hand, the average concentrations of dissolved Fe between the models is much less variable and ranges from 0.35 to 0.81 nmol L-1 (Fig. 1b), or an average of 0.58±0.14 nmol L-1. This apparent constancy reflects an initial view of the ocean Fe cycle in which interior Fe concentrations were held at a quasi-constant value of 0.6 nmol L-1 assuming a constant concentration of Fe-binding ligands (7). Thus the FeMIP models are balancing widely varying Fe input fluxes against relatively constant overall Fe concentrations by tuning the Fe scavenging rate, which is a crucial but poorly known parameter. This results in residence times for Fe that range from <5 to >500 years across the FeMIP models (Fig 1c). This difference is important, as it represents substantial inter-model deviation concerning the timescales over which the different models respond to a perturbation in Fe supply.
When compared statistically against the global dataset, similar levels of variability arise. Some models display correlation coefficients of >0.5, whereas others are slightly anti-correlated. When the FeMIP models are compared against the five GEOTRACES sections, it becomes apparent that those models that represent the newly emerging features of the iron cycle perform much better. For instance, having Fe scavenging rates that vary in space and time, including variable Fe:carbon (C) stoichiometry, multiple Fe sources, and representing ligand concentrations in a dynamic manner, all act to improve the representation of different observed features in the models. Importantly, the IDP2014 provided the opportunity to demonstrate that the issues at hand were specific to Fe, since the models could represent the observed distributions of major nutrients with a much greater degree of skill (5).
The next stage of FeMIP will be a deeper comparison of the processes themselves. Of particular interest is whether GEOTRACES datasets can provide broader assessments of the rates of Fe scavenging – e.g., using other particle-reactive, non-biological tracers such as thorium (8, 9). Equally, the emerging database of GEOTRACES process studies provides an important opportunity to appraise the way different models represent biological iron cycling and in particular, the often-observed importance of recycled and remineralized sources of Fe (10, 11). Finally, the new GEOTRACES intermediate data product 2017 will also facilitate further evaluation of models, providing new section data from the Atlantic, Pacific and Arctic Oceans.
Author
Alessandro Tagliabue (Dept. of Earth, Ocean and Ecological Sciences, School of Environmental Sciences, University of Liverpool)
References
1. L. Bopp et al., Biogeosci. 10(10), 6225-6245, doi:10.5194/bg-10-6225-2013 (2013).
2. C. M. Moore et al., Nature Geosci., doi:10.1038/ngeo1765 (2013).
3. S. Levitus et al., Prog. Oceanogr. 31(3), 245-273, doi:10.1016/0079-6611(93)90003-v (1993).
4. A. Tagliabue et al., Biogeosci. 9(6), 2333-2349, doi:10.5194/bg-9-2333-2012 (2012).
5. A. Tagliabue, A. et al., Glob. Biogeochem. Cycles, doi:10.1002/2015gb005289 (2016).
6. E. Mawji et al., Marine Chem. 177, 1-8. doi:10.1016/j. marchem.2015.04.005 (2015).
7. K. S. Johnson, R. M. Gordon, K. H. Coale, Marine Chem. 57(3-4), 137-161, doi:10.1016/s0304-4203(97)00043-1 (1997).
8. C. T. Hayes et al., Geochim. Cosmochim. Acta 169, 1-16, doi:10.1016/j.gca.2015.07.019 (2015).
9. N. Rogan et al., Geophys. Res. Lett. 43(6), 2732-2740. doi:10.1002/2016gl067905 (2016).
10. P. W. Boyd et al., Glob. Biogeochem. Cycles 29(7), 1028-1043, doi:10.1002/2014gb005014 (2015).
11. R. F. Strzepek et al., Glob. Biogeochem. Cycles 19(4), GB4S26, doi:10.1029/2005gb002490 (2005).

