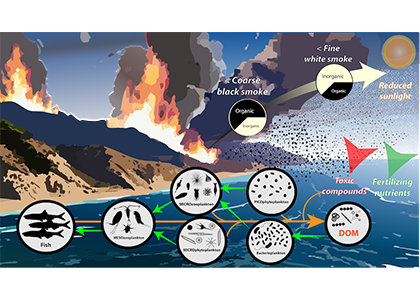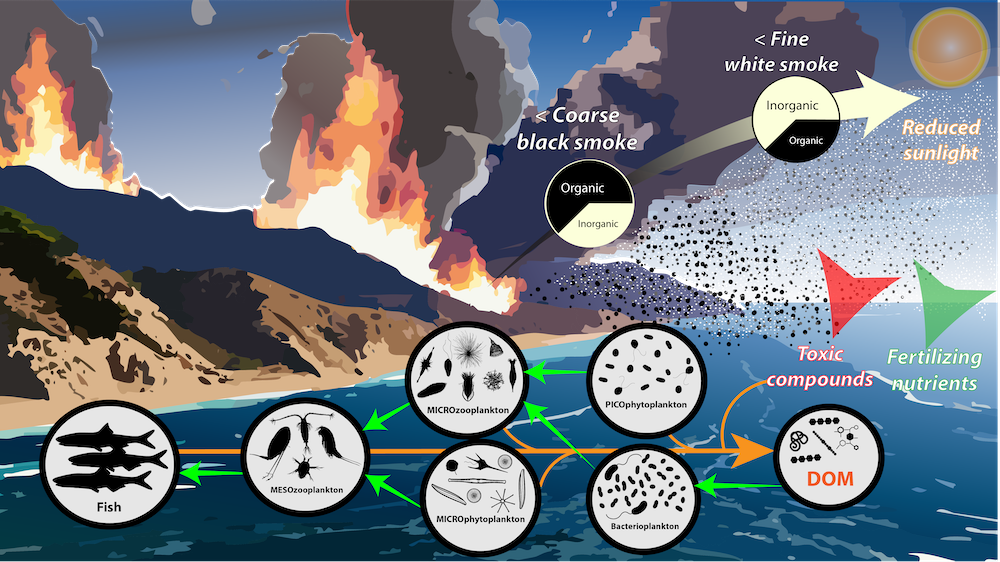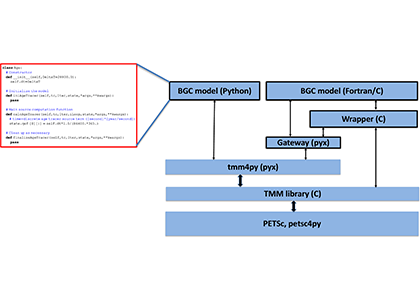
How much organic carbon is actually transported to depth in the Southern Ocean and what are the mechanisms driving seasonal and regional variability? With large-scale remote sensing observations constrained to the surface and the depth-resolved ship-based measurements being scarce, the emergence of the BGC-Argo fleet has opened up a new avenue to explore how carbon is transported from the sunlit surface to the deep ocean.
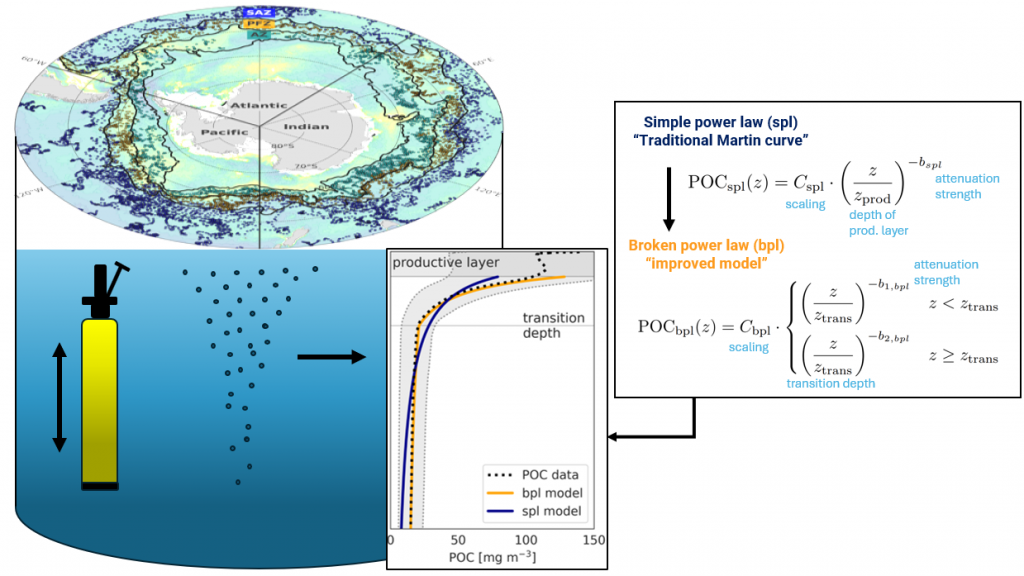
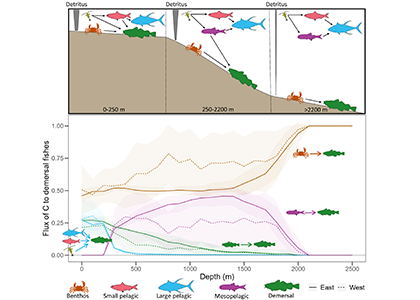
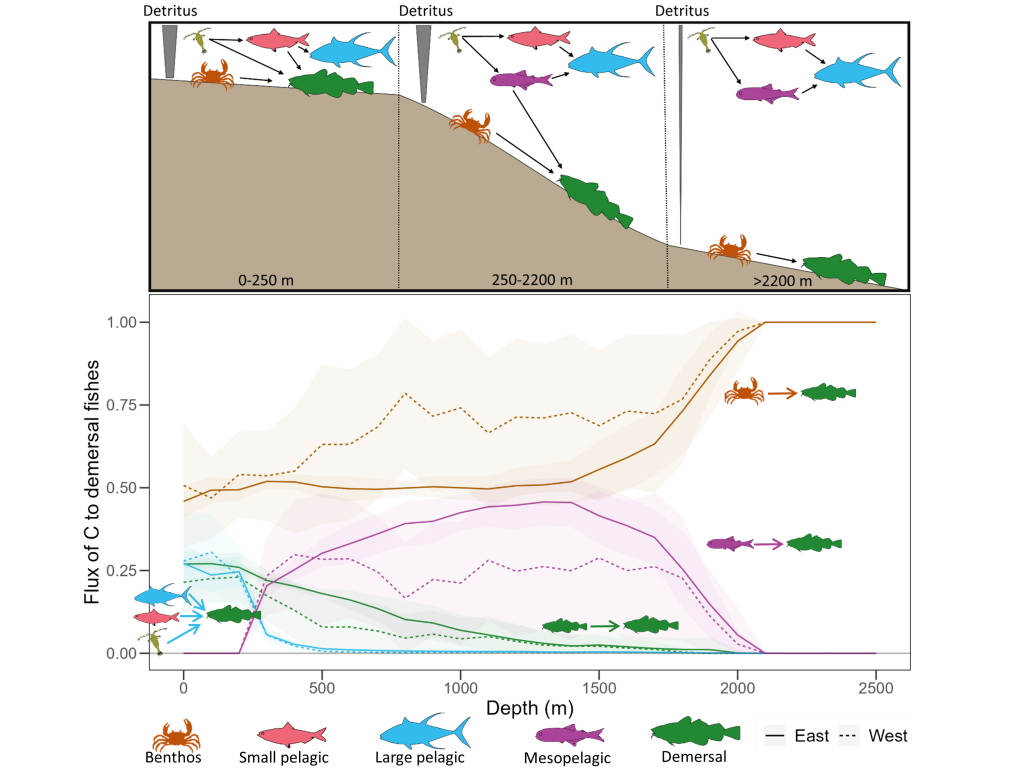
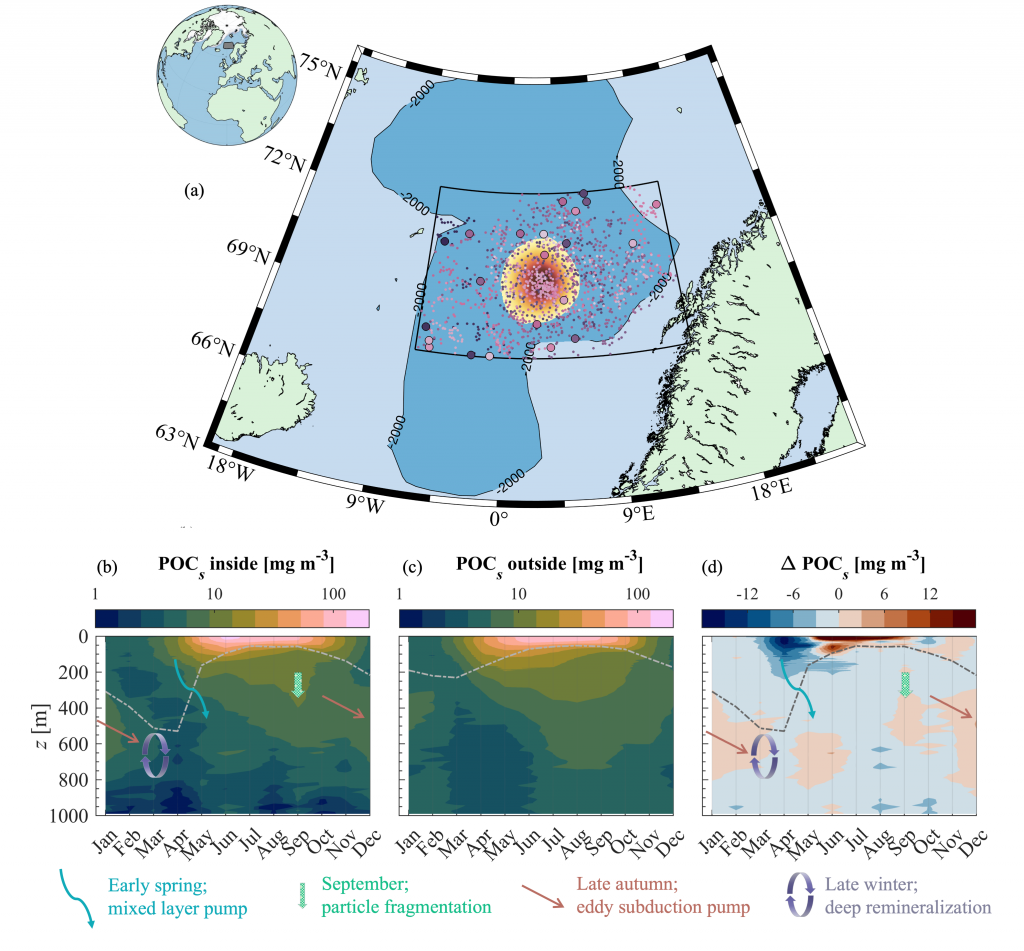
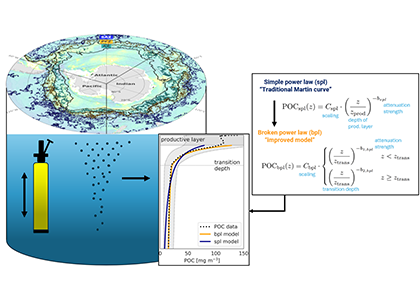
Figure caption: Conceptual illustration of applying a broken power law to POC concentration profiles measured with BGC-Argo floats in the Southern Ocean. The “steepness” or strength of the attenuation is an indicator for how quickly POC attenuates. With the emergence of the BGC-Argo floats, the authors investigate spatial and temporal patterns and possible drivers.
In a recent study published in Communication Earth & Environment, the authors present an improved model to study particle attenuation and discuss what mechanism could explain it: mineral ballasting, zooplankton processes, temperature and net primary production. The work is based on the optical backscatter and chlorophyll-fluorescence observations from the BGC-Argo fleet that are converted into particulate organic carbon (POC) profiles. The float coverage allows us to investigate seasonal patterns and spatial variability across frontal zones and oceanic basins in attenuation strength by applying a power law to the measurements. The traditional version of the simple power law (“Martin curve”) was not able to capture the large variability in the mesopelagic. Instead, we propose a broken power law to better explain higher, observed attenuation coefficients in the upper water column.
This new empirical model for particle attenuation can help guide the experimental studies and model developments needed to help constrain the biological carbon pump and how it may change future emission scenarios.
Authors
Annika Oetjens (University of Tasmania, IMAS, ACEAS)
Tyler Rohr (University of Tasmania, IMAS, AAPP, ACEAS)
Peter Strutton (University of Tasmania, IMAS, ACEAS)
Zanna Chase (University of Tasmania, IMAS, ACEAS)