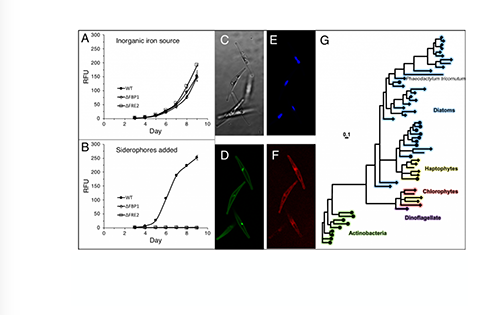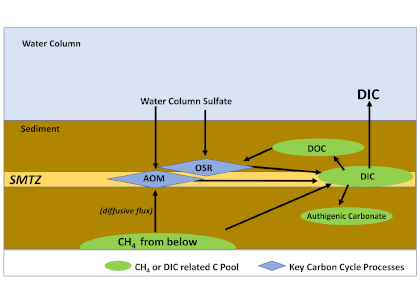
Ocean margins are often characterized by the transport of methane, a potent greenhouse gas, entering from the subsurface and moving towards the seafloor. However, a significant portion of subsurface methane is consumed within shallow sediments via microbial driven anaerobic oxidation of methane (AOM). AOM converts the methane carbon to dissolved inorganic carbon (DIC) and reduces the amount of sulfate that diffuses down from the seafloor towards a sediment interval known as the sulfate-methane transition zone (SMTZ). The SMTZ is where the upward flux of methane encounters the downward diffusive sulfate flux (Figure 1). While the mechanisms of methane production and consumption have been extensively studied, the fate of the DIC that is produced in methane-charged sediments is not well constrained.
In a recent study published in Frontiers in Marine Science, authors used existing reports of methane and sulfate flux values to the SMTZ and synthesized a carbon flow model to quantify the DIC cycling in diffusive methane flux sites globally. They report an annual average of 8.7 Tmol (1 Tmol = 1012 moles) of DIC entering the diffusive methane-charged shallow marine sediments due to sulfate reduction coupled with AOM and organic matter degradation, as well as DIC input from depth (Figure 1). Approximately 75% (average of 6.5 Tmol year–1) of this DIC pool flows upward toward the water column, making it a potential contributor to oceanic CO2 and ocean acidification. Further, an average of 1.7 Tmol year–1 DIC precipitates as methane-derived authigenic carbonates. This synthesis emphasizes the importance of the SMTZ, not only as a methane sink but also an important biogeochemical front for global DIC cycling.
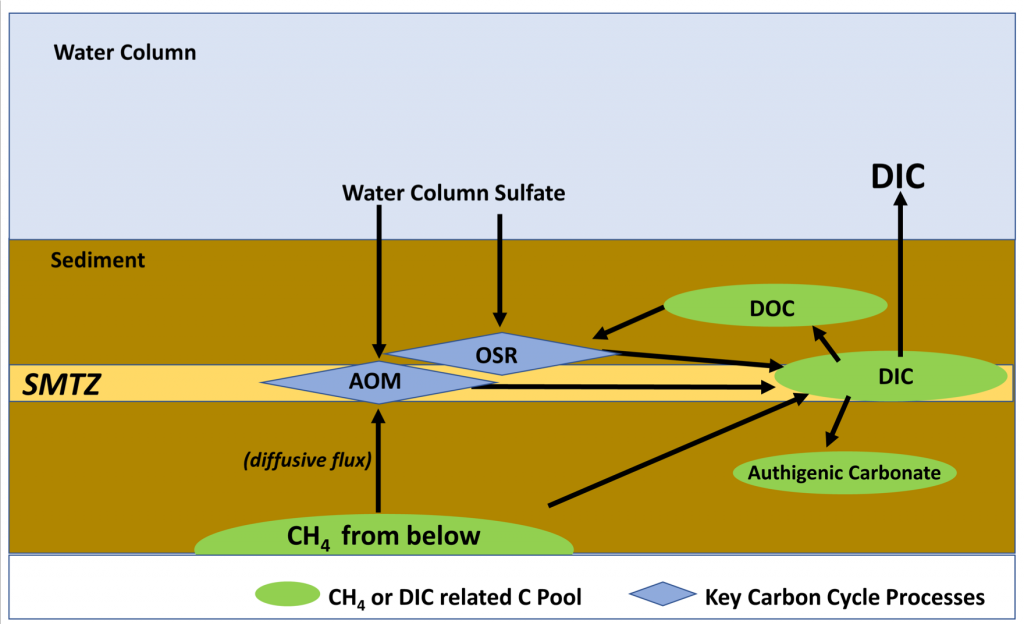
Figure 1: A simplified representation of DIC cycling at diffusive methane charged settings.
The study highlights that regions characterized by diffusive methane fluxes can contribute significantly to the oceanic inorganic carbon pool and sedimentary carbonate accumulation. DIC outflux from the methane-charged sediments is comparable to ~20% global riverine DIC flux to oceans. Methane-derived authigenic carbonate precipitation is comparable to ~15% of carbonate accumulation on continental shelves and in pelagic sediments, respectively. These pathways must be included in coastal and geologic carbon models.
Authors:
Sajjad Akam (Texas A&M University-Corpus Christi)
Richard Coffin (Texas A&M University-Corpus Christi)
Hussain Abdulla (Texas A&M University-Corpus Christi)
Timothy Lyons (University of California, Riverside)