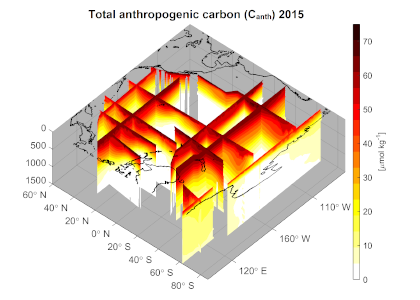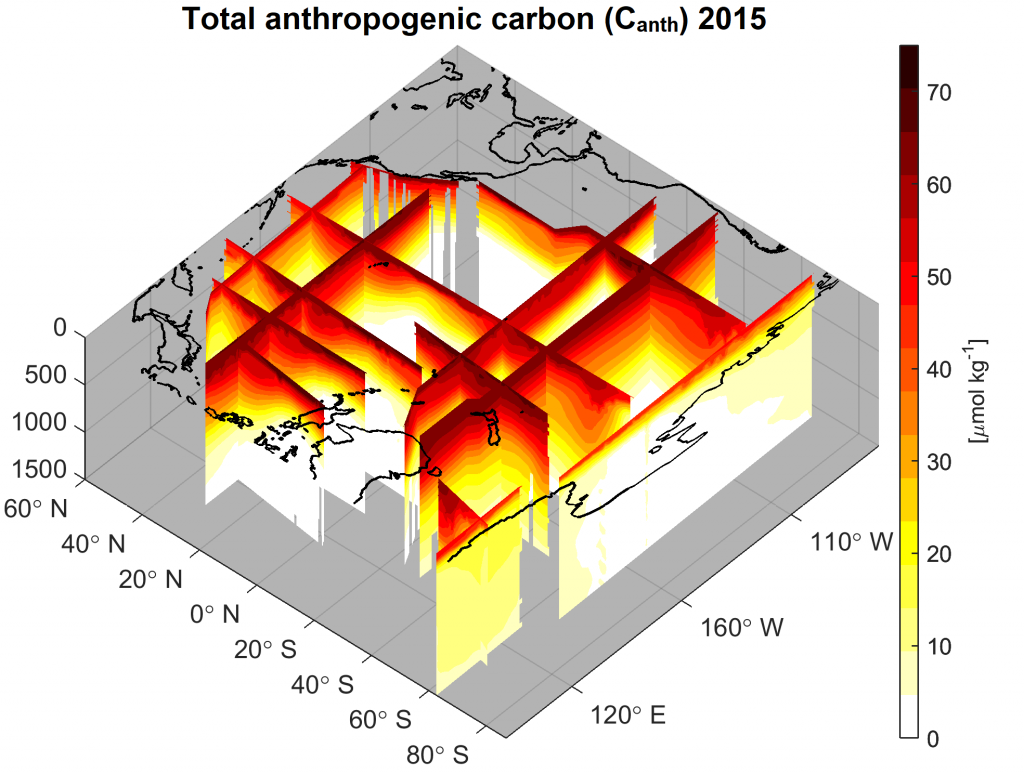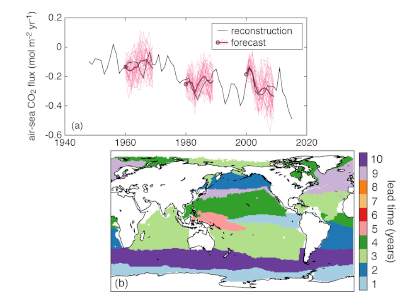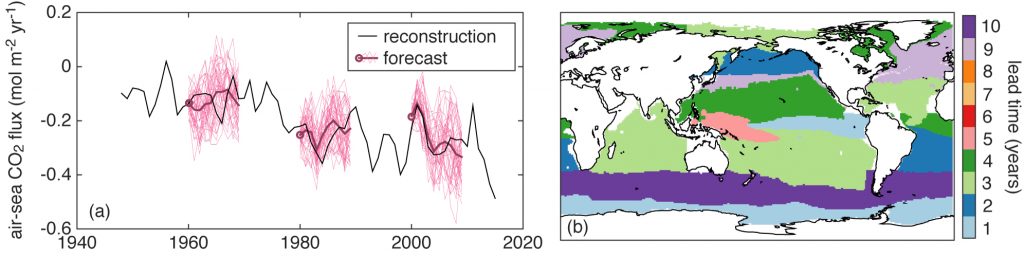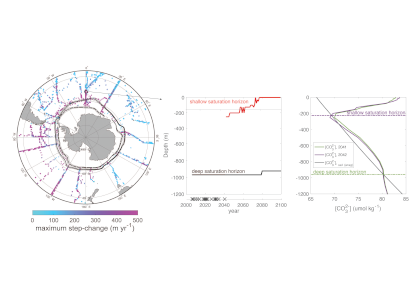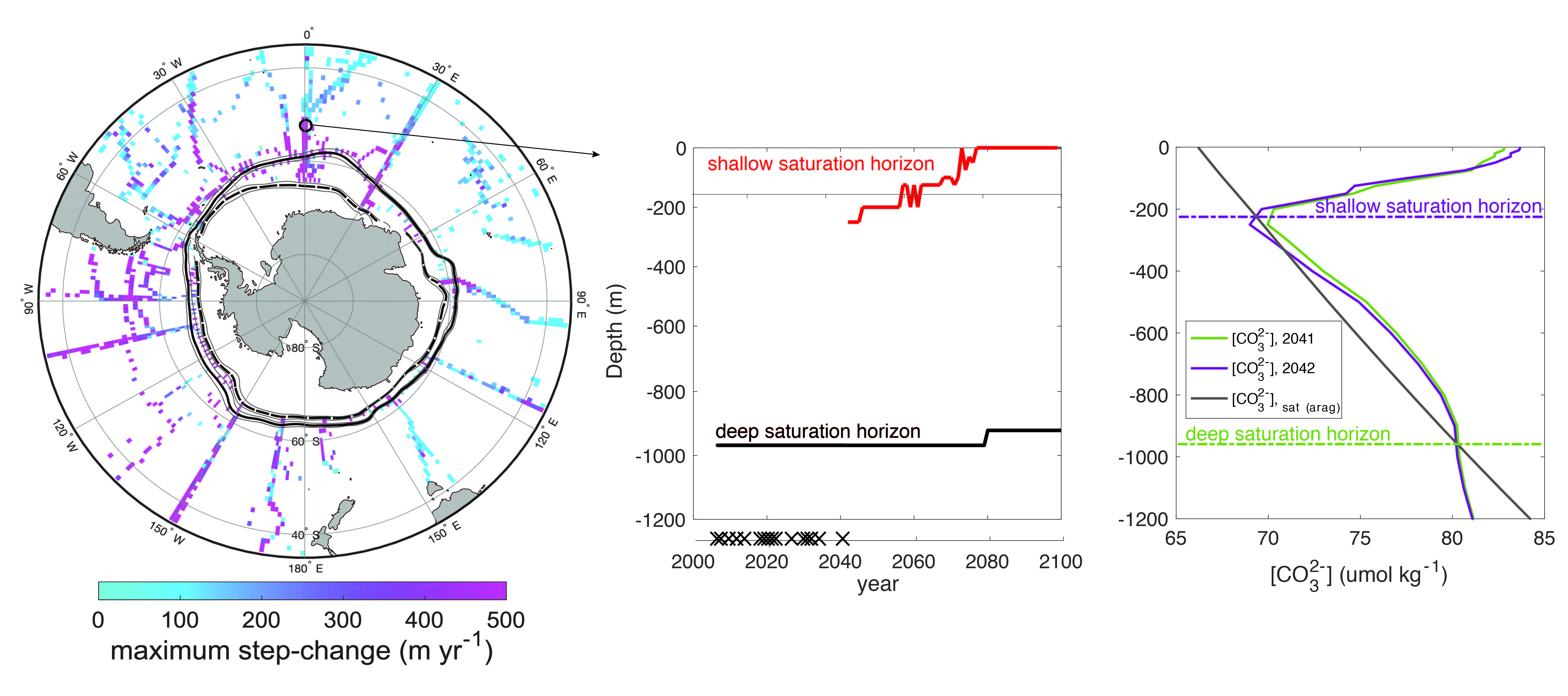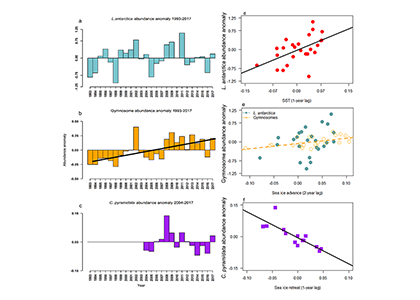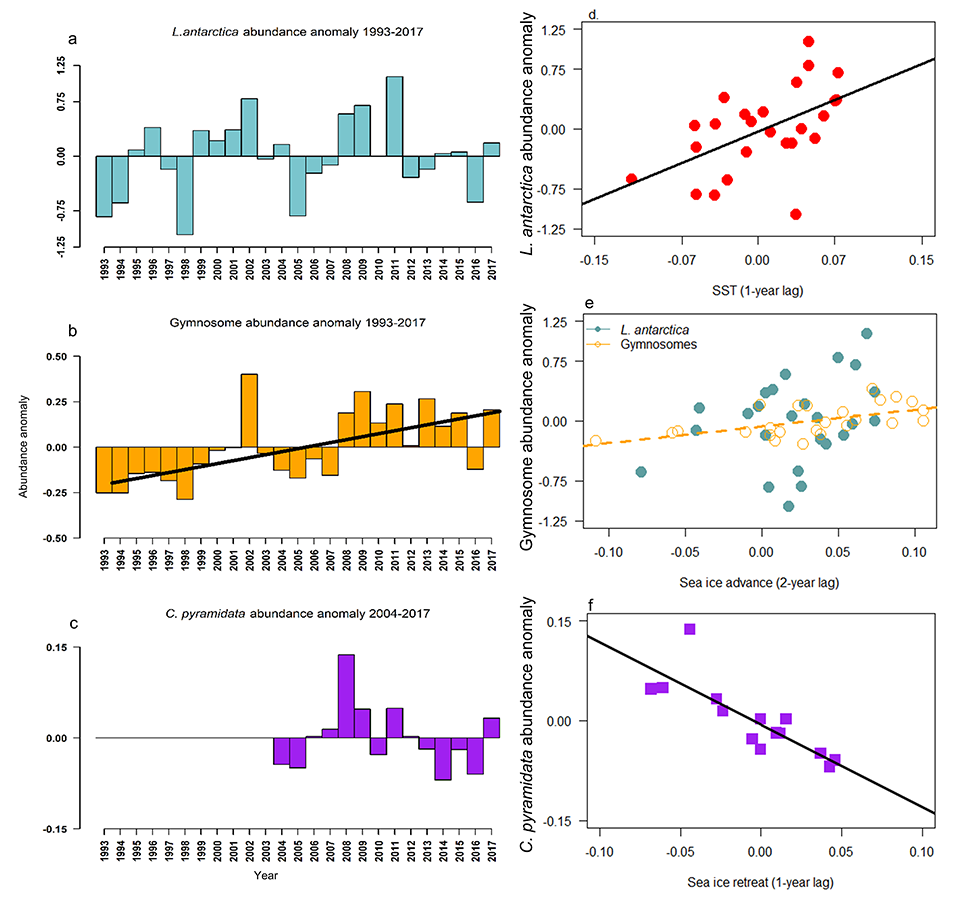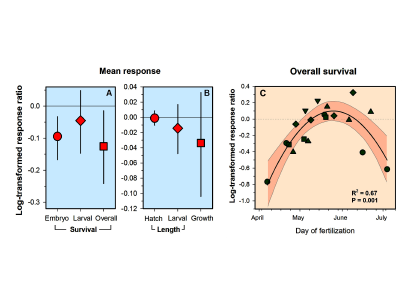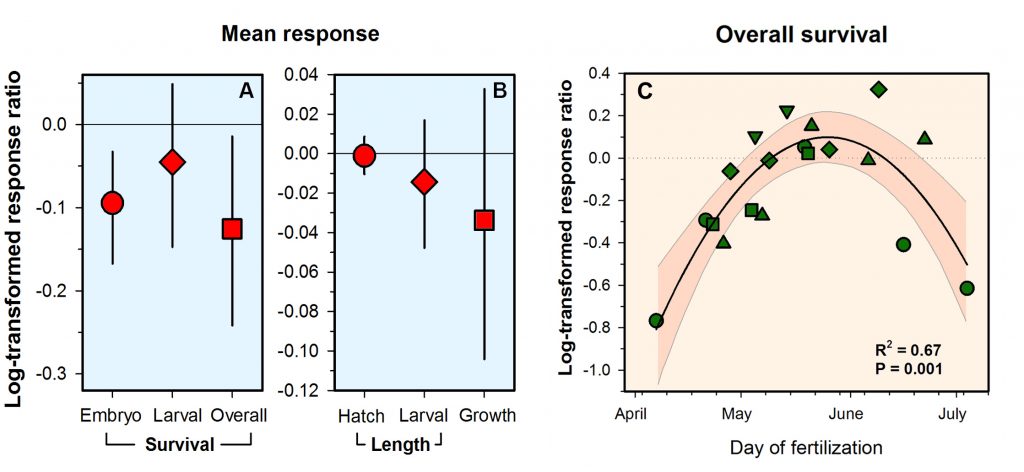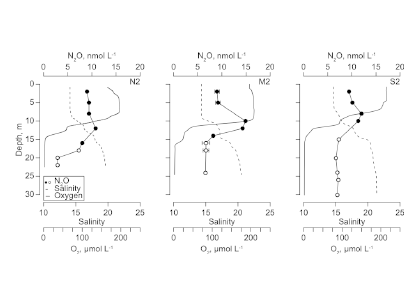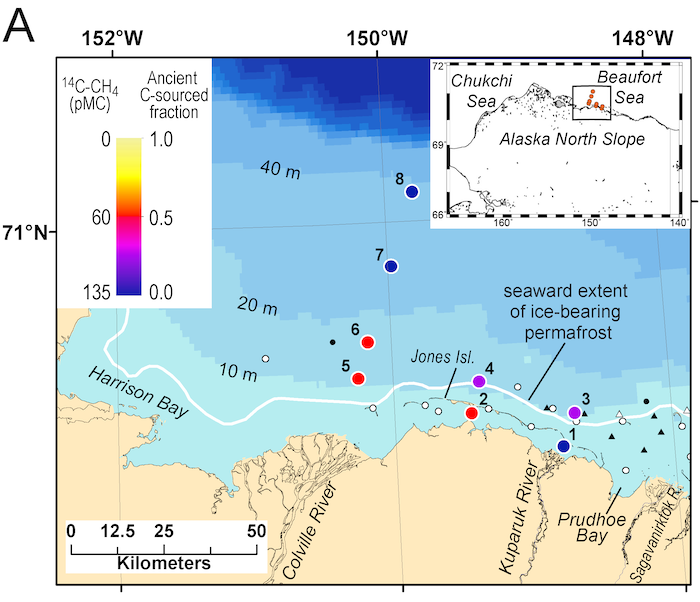
Routine measurements of air-sea gas exchange assume a homogeneous gas concentration across the upper few meters of the ocean. But is this assumption valid? A recent study in Biogeosciences revealed substantial systematic gradients of nitrous oxide (N2O) in the top few meters of the Peruvian upwelling regime. These gradients lead to a 30% overestimate of integrated N2O emissions across the entire region, with local emissions overestimated by as much as 800%.
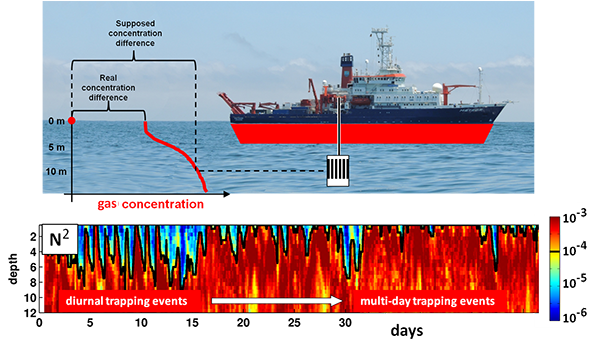
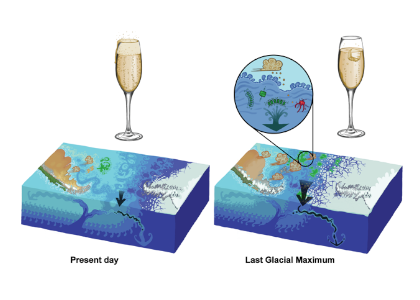
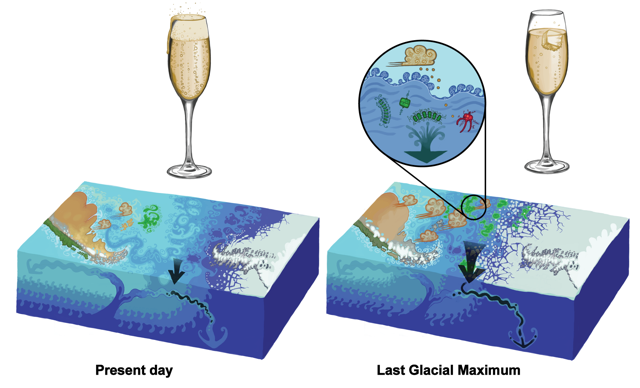
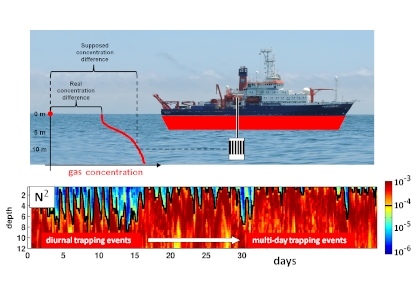
Figure caption: Air-sea gas exchange estimates can be biased by gas concentration gradients within the upper few meters of the ocean; in particular, surface trapping over several days’ duration can generate substantial gradients.
The N2O gradients off Peru form during multi-day events of surface trapping, in which near-surface stratification dampens turbulent mixing. Until now, surface trapping was assumed to be a diurnal (driven by solar warming) process without memory, whereby only weak gradients would form during the hours of trapping and then dissipate. It is likely that multi-day surface trapping occurs in other ocean regions as well. The total impact on emission estimates of different greenhouse gases is yet to be quantified, but given the findings in the Peruvian upwelling system, could be significant globally.
Authors:
Tim Fischer, Annette Kock, Damian L. Arévalo-Martínez, Marcus Dengler, Peter Brandt, Hermann W. Bange (GEOMAR)