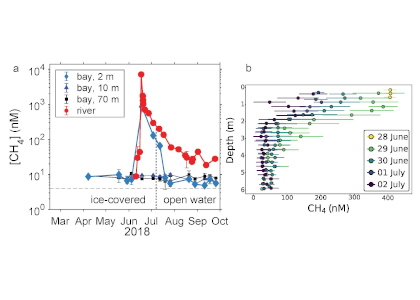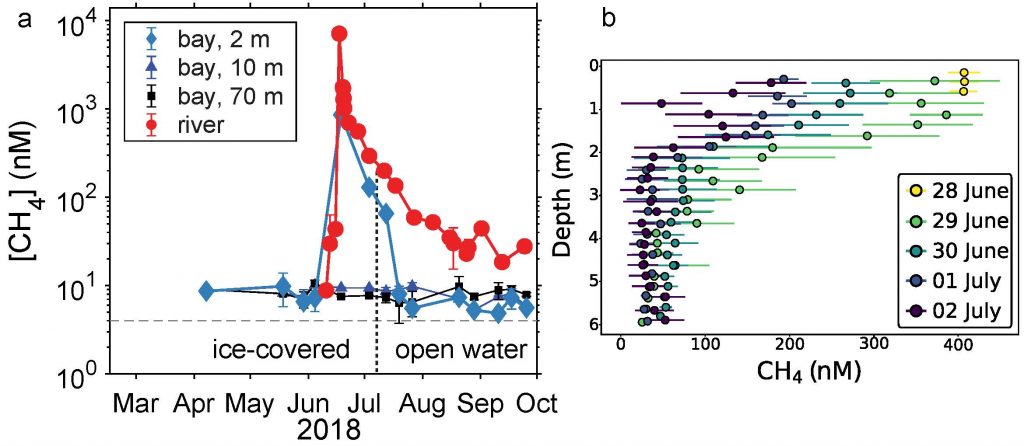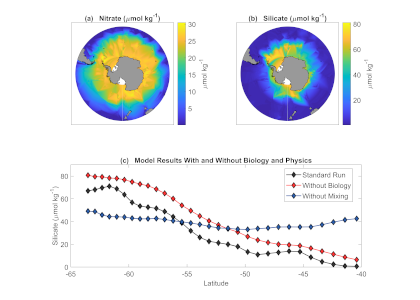
In the Southern Ocean, surface water silicate (SiO4) concentrations decline very quickly relative to nitrate concentrations along a northward gradient toward mode water formation regions on the northern edge (Figure 1a, b). These mode waters play a critical role in driving global nutrient concentrations, setting the biogeochemistry of low- and mid-latitude regions around the globe after they upwell further north. To explain this latitudinal surface gradient, most hypotheses have implicated diatoms, which take up and export silicon as well as nitrogen: (1) Diatoms, including highly-silicified species such as Fragilariopsis kerguelensis, are more abundant in the Southern Ocean than elsewhere; (2) Iron limitation, which is prevalent in the Southern Ocean, elevates the Si:N ratio of diatoms; (3) Mass export of empty diatom frustules pumps silicate but not nitrate to deeper waters.
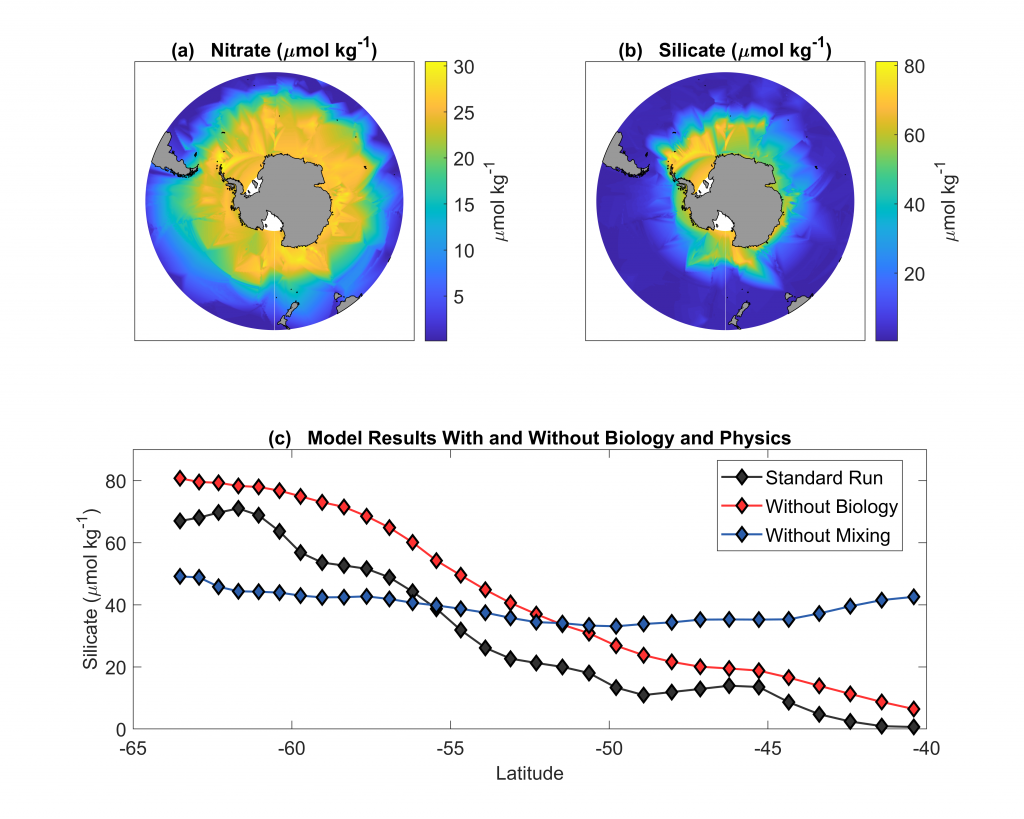
Figure 1: (a) and (b) nitrate and silicate concentrations in surface waters of the Southern Ocean (GLODAPv2_2019 data). (c) Model results of a standard run (black diamonds), a run without biology (red diamonds) and a run without mixing (blue diamonds).
In a recent paper published in Biogeosciences, the authors use an idealized model to explore the relative roles of biological vs. physical processes in driving the observed latitudinal surface nutrient gradients. Over timescales of a few years, removing the effects of biology (no SiO4 uptake or export) from the model elevates silicate concentrations slightly over the entire latitudinal range, but does not remove the strong latitudinal gradient (Figure 1c). However, if the effects of vertical mixing processes such as upwelling and entrainment are removed from the model by eliminating the observed deep [SiO4] gradient, the observed surface nutrient gradient is greatly altered (Figure 1c). These model results suggest that, over short timescales, physics is more important than biology in driving the observed surface water gradient in SiO4:NO3 ratios and forcing silicate depletion of mode waters leaving the Southern Ocean. These findings add to our understanding of Southern Ocean dynamics and the downstream effects on other oceans.
Authors:
P. Demuynck (University of Southampton)
T. Tyrrell (University of Southampton)
A.C. Naveira Garabato (University of Southampton)
C.M. Moore (University of Southampton)
A.P. Martin (National Oceanography Centre)