
Hypoxia is currently one of the greatest threats to coastal and estuarine ecosystems around the world, and this threat is projected to get worse as waters warm and human populations continue to increase. Over the past 35-years, a massive effort has been underway to decrease hypoxia in the Chesapeake Bay by reducing nutrient input from land. Despite this effort, record-breaking precipitation in 2018-2019 fueled particularly large hypoxic volumes in the Bay, calling into question the efficacy of management actions.
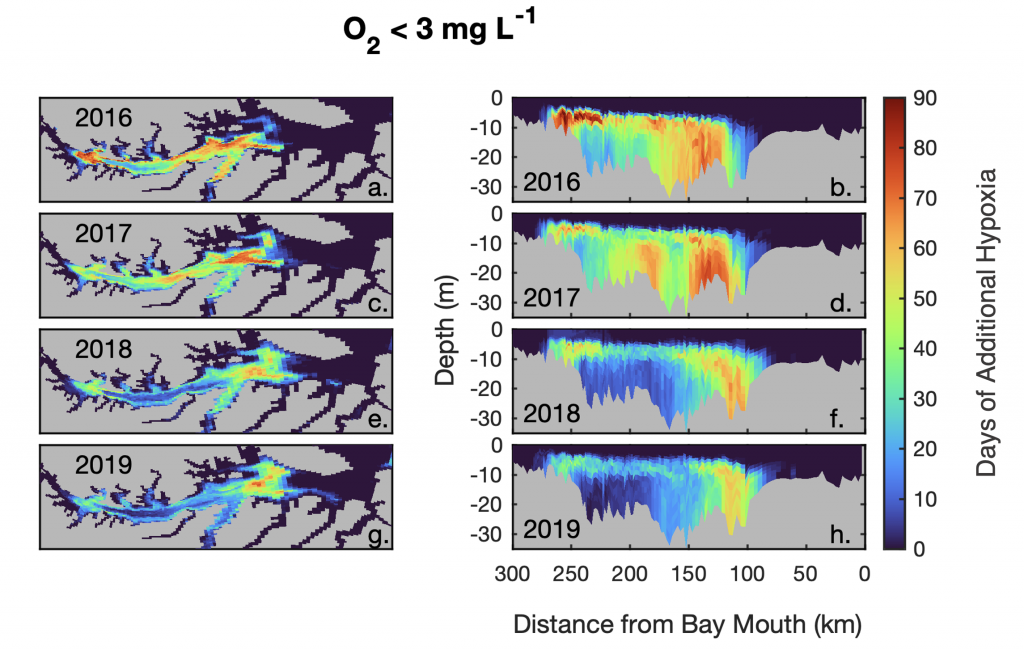
Figure 1. The number of days of additional hypoxia (O2 < 3 mg L-1) that would have occurred in the Chesapeake Bay if the 35 years of nutrient reductions never occurred, as calculated by differences between a realistic numerical model simulation and one with 1985 nitrogen levels. This management effort has had the greatest impact at the northern and southern edges of the hypoxia in the Bay, where there would have been an additional 60-90 days of O2 < 3 mg L-1 if nutrient reductions never occurred.
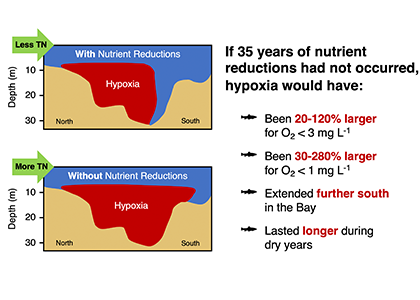
In a recent paper published in Science of the Total Environment, researchers used empirical and numerical modeling to quantify the impact of nutrient management efforts on hypoxia in the Chesapeake Bay. Results suggest that if the nutrient reduction efforts beginning in 1985 had not taken place, hypoxia would have been ~50–120% greater during the average discharge years of 2016–2017 and ~20–50% greater during the wet years of 2018–2019. The management impact was most pronounced in regions of the Bay where the hypoxia season would have been 60-90 days longer if nutrient reductions did not occur (Figure 1).
Although these results suggest that management has reduced hypoxic conditions in the Bay, additional analysis revealed that warming temperatures have already offset 6-34% of this improvement. This highlights the importance of factoring in climate change when setting future management goals.
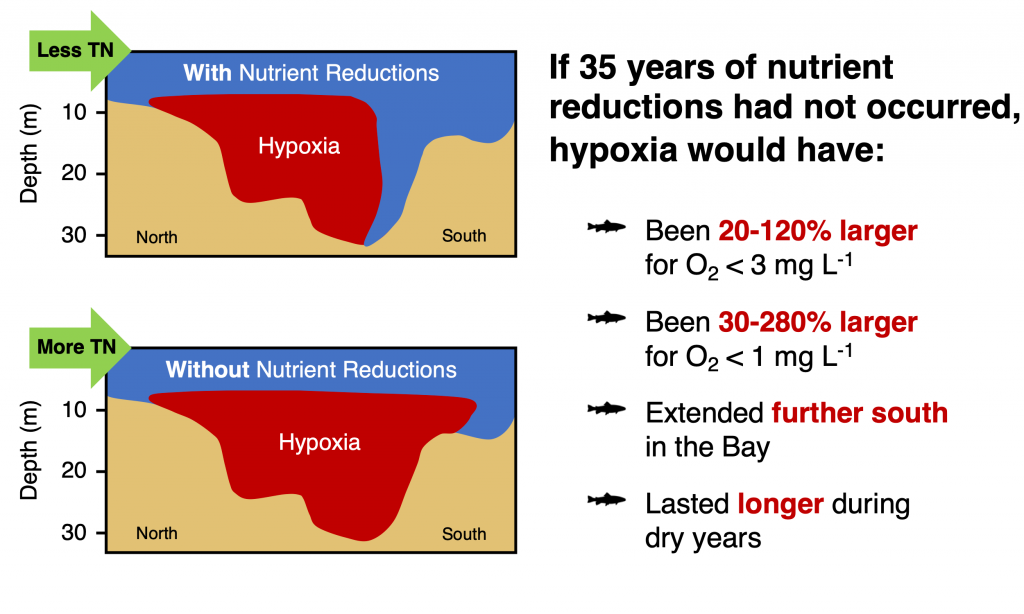
Figure 2. The number of days of additional hypoxia (O2 < 3 mg L-1) that would have occurred in the Chesapeake Bay if the 35 years of nutrient reductions never occurred, as calculated by differences between a realistic numerical model simulation and one with 1985 nitrogen levels. This management effort has had the greatest impact at the northern and southern edges of the hypoxia in the Bay, where there would have been an additional 60-90 days of O2 < 3 mg L-1 if nutrient reductions never occurred.
Authors:
Luke T. Frankel (Virginia Institute of Marine Science, William & Mary)
Marjorie A. M. Friedrichs (Virginia Institute of Marine Science, William & Mary)
Pierre St-Laurent (Virginia Institute of Marine Science, William & Mary)
Aaron J. Bever (Anchor QEA)
Romuald N. Lipcius (Virginia Institute of Marine Science, William & Mary)
Gopal Bhatt (Pennsylvania State University; Chesapeake Bay Program)
Gary W. Shenk (USGS; Chesapeake Bay Program)

