
Oxygen Deficient Zones (ODZs) are naturally occurring functionally anoxic regions of the open ocean which can act as proxies for early Earth’s anoxic ocean. Without free oxygen, microorganisms in these regions use alternative electron acceptors to oxidize organic material. These functionally anoxic regions are also hotspots for chemoautotrophic pathways. Some microorganisms can use arsenic based compounds to oxidize organic material, and others can couple nitrate reduction with arsenic oxidation supporting autotrophic carbon fixation thus linking arsenic respiration with carbon and nitrogen cycling. While arsenic concentrations in modern oceans are relatively low, the Precambrian ocean likely had periods of high arsenic concentrations. Integrating over time and space of anoxic waters, arsenic-based metabolisms may have had significant implications for the biogeochemical cycling of not only arsenic, but also carbon and nitrogen.
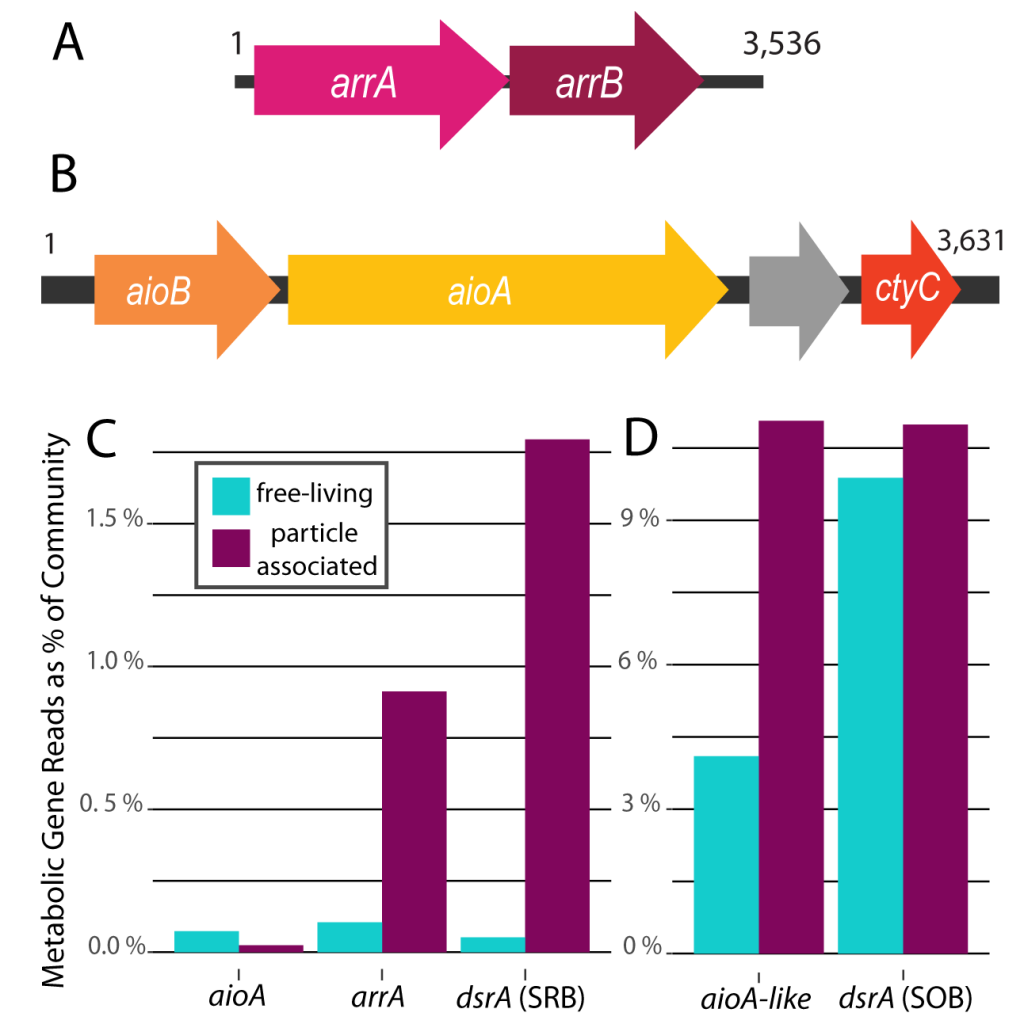
Figure 1: Arsenotrophic genes identified in the Eastern Tropical North Pacific Oxygen Deficient Zone. (A) Genomic complement for dissimilatory arsenate reduction assembled from metagenomes which likely supports respiration of organic matter. (B) Genomic complement for putative chemoautotrophic arsenite oxidation pathway assembled from metagenomes which may couple with nitrate reduction to support organic matter production. (C) Relative abundance of genes associated with arsenite oxidase (aioA), dissimilatory arsenate reduction (arrA), and forward dissimilatory sulfite reductase (dsrA) associated with sulfur reduction; abundance shown as a relative contribution to the total microbial community as estimated by abundance of RNA polymerase genes (rpoB). The genes arrA and forward-dsrA are more abundant in the particulate fraction, whereas aioA is more abundant in the free-living fraction. (D) Relative abundance of genes in the microbial community for the more abundant genes aioA-like and reverse form of dsrA associated with sulfur oxidation. aioA-like genes are relatively more abundant within the particulate fraction, with no strong partitioning between fractions identified for the reverse-dsrA genes. Arsenical reduction and chemoautotrophic arsenical oxidation are likely performed by different microbial groups within the ODZ communities.
Recent work in PNAS identified gene sequences for a complete arsenic respiratory cycle from Eastern Tropical North Pacific (ETNP) ODZ metagenomes. The authors identified arsenotrophic genes for dissimilatory arsenate reduction from one group of microorganisms and genes for a putative chemoautotrophic arsenite oxidation pathway from another group within the ETNP ODZ microbial community. Analysis of genomic sequences from a free-living sample and from particulate-associated sample indicate niche differentiation of these pathways—arsenate reduction genes enriched within the particulate fraction and arenite oxidation enriched in the free-living water column. In addition to the presence of these genes in metagenomes, the authors identified the active expression of these arsenotrophic genes in publicly available metatranscriptomes from the ETNP and Eastern Tropical South Pacific ODZs. Theyalso found an abundance of sequences in the ETNP ODZ for the gene aioA-like, which is a closely related enzyme to arsenite oxidase (aioA), but with an unconfirmed function. The identification of these actively expressed genes in modern ODZs enables further investigation of these cycles that were likely important in early oceans. These findings also highlight that there are still yet to be discovered respiratory pathways in ODZs. Arsenotrophy, in conjunction with other niche respiratory pathways – both known and as yet undiscovered – likely sum to a considerable contribution of energy flow and elemental cycling through these anoxic systems.
Authors:
Jaclyn Saunders (University of Washington; present affiliation Woods Hole Oceanographic Institution)
Clara Fuchsman (University of Washington; present affiliation Horn Point Laboratory)
Cedar McKay & Gabrielle Rocap (University of Washington)
See related University of Washington press-release

